Мохообразные представляют собой единственный отдел среди высших растений, объединяющий растительные организмы, у которых гаметофит является преобладающим поколением. Собственно то растение, которое мы представляем себе при слове «мох», и является гаметофитом (невзрачный спорофит неспециалисты, как правило, вообще оставляют без внимания). Следует отметить, что, несмотря на чрезвычайную распространенность этого слова, называть всех представителей отдела мохообразных просто мхами неправильно, поскольку этот отдел также включает в себя растения, которые в более строгой номенклатурной системе называются иначе.
Мохообразные характеризуются относительной простотой строения. Как и подобает любому высшему растению, тело мохообразных расчленено на вегетативные органы - стебель и лист (рис. 186, 187), у наиболее простых тело представляет собой слоевище (таллом), уплощенное в дорзо-вентральном направлении (рис. 188). Однако у мохообразных все вегетативные органы не могут считаться истинными, поскольку они принадлежат гаметофиту и не гомологичны настоящим органам спорофита. Поэтому ученые-бриологи (бриология — наука, изучающая мохообразные) именуют их «стебли» и «листья» каулидиями и филлидиями соответственно. Мохообразные не имеют корней и к субстрату прикрепляются нитевидными ризоидами, которые у различных форм могут быть одно- или многоклеточными. Поглощение воды и других необходимых веществ растением осуществляется не только ризоидами, но и через всю поверхность тела, чему в значительной мере способствует полное (или почти полное) отсутствие на поверхности защитной кутикулы. Это обстоятельство привносит в жизнь мохообразных немало дополнительных трудностей, поскольку вода не только с легкостью поступает внутрь растения, но и также легко выходит из него, что вынуждает их обживать места с повышенной влажностью. Однако, несмотря на это, некоторые ксерофитные мохообразные приспособились к засушливым условиям.

Рис. 186. Листостебельные мхи (верхоплодные):
1 - тортула полевая (Tortula ruralis); 2 - сплахнум красный (Spiachnum mbrum): 3 - сплахнум желтый (S. luteum); 4 - буксбаумия безлистная (Buxbaumia aphylla); 5 - родобриум розетковидный (Rhodobryum roseum); 6 - дикранум метловидный (Dicranum scoparium); 7 - андрея скальная (Andreaea rupestris); 8 - мниум волнистый (Mnium undulatum); 9 - лолитрихум обыкновенный, или кукушкин лен (Polytrichum commune); 10 - ракомитриум шерстистый (Rhacomitrium lanuginosum); 11 - леукобриум сизый (Leucobryum glaucum); 12 - дикранум многоножковый (Dicranum polysetum); 13 - полия поникшая (Pohlia nutans); 14 - фунария зигромвтрическая (Funaria hygrometrica); 15 - аулакомниум вздутый (Aulacomnium turgidum); 16- папуделпа оттопыренная (Paludella squamosa) (по «Жизнь растений», т. 4)
При этом они способны практически полностью обезвоживаться (разумеется, в этом периоде растение находится в состоянии покоя и физиологические функции в нем временно приостанавливаются), но при последующем увлажнении активно впитывают воду и быстро возвращаются к нормальной жизни.

Рис. 187. Листостебельные мхи (бокоплодные):
1 - фонтиналис противопожарный (Fontinalis antipyretica); 2 - туидиум тамарисковидный (Thuidium tamariscinum); 3 - миуроклада Максимовича (Myuroclada maximoviczii); 4 - калливргон лозовидный (Calliergon sarmentosum); 5 - неккера курчавая (Neckera crispa); 6 - леукодон повислый (Leucodon pendulus); 7 - климациум японский (Climacium japonicum); 8 - ортотециум золотистый (Orthothecium chryseum); 9 - дрепанокладус отвернутый (Drepanocladus revolvens); 10- птилиум гребенчатый, или страусово перо (Ptilium crista-castrensis) (по «Жизнь растений», т. 4)
Механизм такого физиологического поведения до конца еще не ясен.
Внутренний транспорт поглощенной воды и растворенных в ней минеральных веществ затруднен, ввиду полного отсутствия проводящей ткани у большинства мохообразных (лишь у немногих из них имеются слабо развитые проводящие структуры, напоминающие элементы флоэмы и ксилемы). Кроме проводящих, у мохообразных отсутствуют или слабо развиты и большинство других тканей, в частности механические, что существенно ограничивает размеры растений. Поэтому наземные формы имеют весьма скромные габариты - до нескольких сантиметров. Водные мохообразные используют воду в качестве дополнительной опоры и нередко превышают полметра в длину.

Рис. 188. Печеночники и антоцеротовые:
1 - маршанция многообразная (Marchantia polymorphe); 2 - пеппия эпифилльная (Pellia epiphylla); 3 - антоцерос точечный (Anthoceros punctatus); 4 - риччия Фроста (Riccia frostii); 5 - риччиокарпус плавающий (Ricciocarpus natans) (по «Жизнь растений», т. 4)
Анатомическое строение мохообразных очень разнообразно. В самом простом варианте все растение может состоять из похожих друг на друга клеток (они различаются лишь по толщине стенок). У более сложных мхов клетки подвергаются некоторой дифференцировке. Например, у печеночного мха маршанции клетки слоевища разделены на два слоя: верхний ассимиляционный и нижний - основной (рис. 189). Клетки ассимиляционного слоя содержат хлоропласты и фотосинтезируют, тогда как клетки основного слоя их лишены.

Рис. 189. Маршанция:
I - поперечный разрез таллома:
А - при малом увеличении;
В - при большом (1 - амфигастрии, 2 - ризоиды, 3 - ассимиляторы, 4 - устьице);
II - ризоиды: А - простой; Б, В-язычковые;
III - поверхностный срез (сверху) таллома (4 - устьице, 5 - воздушная камера) (по Н. А. Комарницкому и соавт., с изменениями)
В верхней эпидерме находятся устьица, а в нижней имеются однослойные чешуйки - амфигастрии – и ризоиды, которые могут быть простыми и язычковыми (они имеют выросты внутри).
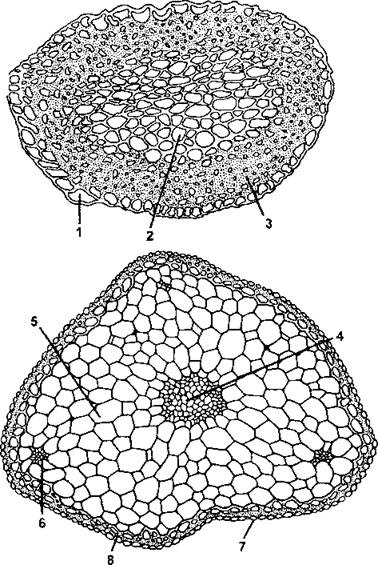
Рис. 190. Стебли бриевых на поперечном срезе.
Вверху - пилотрихидиум (Pilotrichidium antillarum):
1 - гиалодерма; 2 - основная ткань; 3 — кора.
Внизу - мниум (Mnium dnclidioides): 4 - центральный пучок; 5 - основная ткань; 6 - листовой след; 7 - гиалодерма; 8 - кора (по И. И. Абрамову и соавт.)
У сложно организованных листостебельных мхов расположенная снаружи кора содержит вытянутые клетки с утолщенными стенками - стереиды, которые выполняют опорную функцию (эту ткань еще называют склеродермой). У некоторых мхов снаружи имеется слой клеток без хло- ропластов, но с тонкими прозрачными стенками - гиалодерма (или наружная кора). Клетки гиалодермы способны впитывать воду и передавать ее внутренним тканям. Под корой располагаются паренхимные клетки основной ткани, в которых находятся хлоропласты и откладываются запасные вещества (рис. 190). В основной ткани можно обнаружить проводящие пучки и листовые следы (некоторые из них доходят до центрального пучка, а другие слепо теряются в паренхиме). За редким исключением, мохообразные - многолетние растения.
Развитие мохообразных не отличается от других высших растений и представляет собой чередование гаплоидной и диплоидной фаз развития. Преобладающим поколением является гаметофит. Он развивается из споры, ядро которой имеет гаплоидный набор хромосом, поэтому все клетки гаметофита тоже гаплоидные. Первоначально из споры развивается протонема (греч. protos - первый и пета - нить), имеющая вид ветвящейся нитчатой водоросли или пластинки (рис. 191). Никакого запаса питательных веществ спора не содержит, поэтому молодой гаметофит должен их синтезировать самостоятельно посредством фотосинтеза. Наличие необходимой для этого хлоренхимы определяет зеленую окраску гаметофита. Дальнейшее развитие растения из протонемы зависит от его систематического положения.
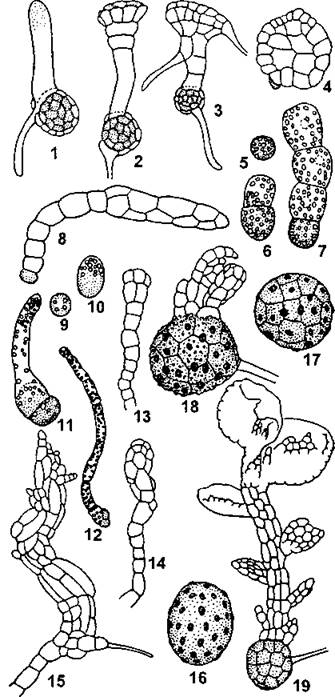
Рис. 191. Типы прорастания спор и развития молодых растений:
1 —4 - у маршанциевых (Reboulia hemisphaerica); 5 — 8-у метцгериевых (Riccardia miyakeana); 9 - 15 — у юнгерманниевых (Plectocolea radicellata); 16 - 19-у фрулланиееых (Frullania tnincabfolia) (по И. И. Абрамову и соавт.)
У печеночных мхов многократное деление верхушечной клетки в трех плоскостях дает начало пластинчатым структурам, на которых впоследствии разовьются половые органы, в связи с чем они получили название гаметофоров (рис. 192, 193). Более сложное формирование гаметофита происходит у лиственных мхов. Их гаметофоры имеют вид облиственных побегов, и развиваются они из почек, которые формируются на протонеме. Поскольку почки на протонеме закладываются достаточно кучно, это приводит к образованию пучков гаметофоров, что и бросается в глаза при встрече с мхами в их естественной среде обитания.
На гаметофорах образуются половые органы - женские архегонии и мужские антеридии. Чаще всего на одном растении развиваются органы только одного пола - однодомные мхи, но нередко имеет место двудомность (когда на одной особи формируются как женские половые органы, так и мужские). Наконец у отдельных форм отмечается многодомность. В этом случае на одном растении образуются как однополые гаметофоры, так и обоеполые. Архегонии и антеридии обычно располагаются группами и в типичном случае окружены различного рода защитными образованиями. Чаще всего с помощью подставок они возвышаются над поверхностью гаметофита (см. рис. 188-1), но нередко погружаются в его глубину. Среди развитых гаметангиев часто встречаются недоразвитые стерильные структуры - парафизы.
Антеридии представляют собой овальные тельца, окруженные тонкой однослойной оболочкой (см. рис. 192). Они наполнены сперматогенными клетками, которые при митотическом делении дают начало двум подвижным сперматозоидам, снабженным двумя жгутиками. Напомним, что клетки гаметофита изначально гаплоидные, поэтому половые клетки образуются не в результате мейоза, как это обычно имеет место у диплоидных организмов, а посредством митоза.
Архегоний устроен сложнее и обычно представляет собой структуру, по своей форме напоминающую бутылку (см. рис. 192-Г-5; рис. 193). В утолщенной его части, называемой брюшком, находится крупная яйцеклетка, которая также образуется в результате митоза. Внутри суженной шейки в один ряд располагаются шейковые клетки, одна из которых - брюшная шейковая клетка - находится над яйцеклеткой.

Рис. 192. Половые органы мохообразных:
А - маршанция обыкновенная (Marchanda polymorpha): 1 - продольный разрез мужской подставки с антеридиями; 2 - изолированный антеридий; 3 - ножка антеридия; 4 - двужгутиковый сперматозоид;
Б - антеридии калобриума (Calobrium);
В - антеридии андреева мха (Andreaea rupestris);
Г - радула (Radula): 5 - антеридий; 6 - архегонии (по Н. А. Комарницкому и соавт., с изменениями)

Рис. 193. Архегонии мохообразных:
А - продольный разрез женской подставки с архегониями маршанции обыкновенной (Marchanda polymorphe):
Б - архегонии калобриума (Calobrium) (по Н.А. Комарницкому и соавт.)
Несмотря на то обстоятельство, что мохообразные представляют собой наземные растения, оплодотворение у них возможно только в присутствии капельножидкой воды. Учитывая небольшие размеры мхов, им для оплодотворения необходимо очень немного влаги, что во многом облегчает им ситуацию. Эффективность процесса обеспечивается подвижностью снабженных жгутиками сперматозоидов. Полагают, что их привлекают вещества, выделяемые канальцевыми клетками архегония, которые к тому моменту ослизняются. Через шейку сперматозоиды проникают в брюшко архегония и оплодотворяют находящуюся там яйцеклетку. В результате образуется диплоидная зигота, которая по прошествии определенного периода покоя дает начало диплоидному поколению - спорофиту.
Спорофит мохообразных называется спорогон (греч. spora - семя и gone - рождение) и среди всех высших растений устроен наиболее просто. В типичном случае он представляет собой коробочку, представляющую собой спорангий, которая посредством ножки переходит в тело гаметофита (см. рис. 186). Разросшаяся и видоизмененная верхняя стенка архегония прикрывает коробочку и называется колпачком, или калиптрой (рис. 194). Не обладая зеленой окраской, спорогон современных мохообразных не содержит хлорофилла и не способен самостоятельно снабжать себя органическими соединениями.
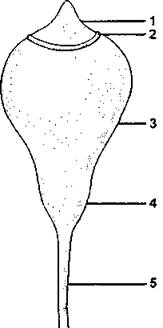
Рис. 194. Строение коробочки листостебельных мхов (схема):
1 - крышечка: 2 - колечко; 3 - урночка: 4 - шейка; 5 - ножка (по И. И. Абрамову и coaвт.)
Поэтому все нужные для развития вещества спорогон получает от гаметофита посредством проникающей в его ткани нижней расширенной части ножки - гаустории. Внутри коробочки многочисленные материнские клетки спор делятся мейозом и дают начало тетрадам гаплоидных спор, посредством которых осуществляется бесполое размножение мохообразных (рис. 195). У печеночных мхов в коробочке среди спор находятся особые клетки, которые видоизменяются в пружинки - элатеры. Они обладают гигроскопичностью, т. е. способностью втягивать в себя пары воды при повышении влажности атмосферного воздуха. При этом элатеры раскручиваются, а при понижении влажности вновь скручиваются, перемешивая и разрыхляя при этом массу спор, что способствует выталкиванию их из коробочки после того, как она раскрывается несколькими створками.

Рис. 195. Развитие спор в спорогоне маршанции:
I - молодой спорогон: 1 - разросшийся перианций, 2 - брюшко архегония, 3 - коробочка, 4 - короткая ножка, 5 - гаустория; II - часть более старого спорогона (видны споры и пружинки); III - материнские клетки спор: 6 - два ряда материнских клеток спор и часть пружинки (удлиненная клетка), 7 - материнские клетки спор делятся с образованием тетрад, 8 - споры и пружинка (по Н. А. Комарницкому и соавт.)
Лиственные мхи элатерами не обладают. Проблема равномерного рассеивания спор у них решается благодаря особой структуре - перистому, который имеется у всех лиственных мхов, за исключением сфагновых и андреевых мхов (рис. 196). Перистом представляет собой множество мелких зубчиков, расположенных в один или несколько рядов вокруг расширенной части коробочки (урночки).

Рис. 196. Типы перистомов у листостебельных мхов:
1 - тиммия (Timmia); 2 - октоблефарум (Octoblepharum); 3 - улота (Ulota); 4 - фонтиналис (Fontinalis); 5 - тейлория (Tayloria); 6 - цинклидиум (Ginclidium) (по И. И. Абрамову и соавт.)
Выше располагается прикрытая крышечкой шейка, которая отграничивается от урночки кольцом, по уровню которого она и отделяется после засыхания коробочки. Подобно элатерам, зубцы перистома гигроскопичны. Во влажную погоду они насыщаются водой, что приводит к их деформации и перекрытию отверстий в коробочке. Споры при этом не выбрасываются, но и вода не может попасть в коробочку. В сухую погоду происходит обратный процесс. Зубцы перистома высыхают и отгибаются наружу, открывая тем самым отверстия в коробочке, и споры, выпадающие из нее, подхватываются ветром и переносятся на расстояния нередко значительно отдаленные от материнского растения. Громадное большинство спор гибнет, попадая в неблагоприятную среду, но спор производится так много, что часть из них обязательно оказывается во влажной почве и там из них прорастает протонема, давая начало молодому гаметофиту. Схема жизненного цикла печеночного мха пеллии показана на рис. 197.

Рис. 197. Схема жизненного цикла печеночного мха пеллии (Реlliа) (по Н. Грину и соавт., с изменениями)
Наряду с описанным выше циклом развития со сменой гаметофитного и спорофитного поколений у мохообразных также широко распространено вегетативное размножение. У печеночников образуются выводковые тельца и выводковые корзиночки (рис. 198), а у листостебельных мхов почти из всех структур гаметофита могут образовываться вторичные протонемы (рис. 199).
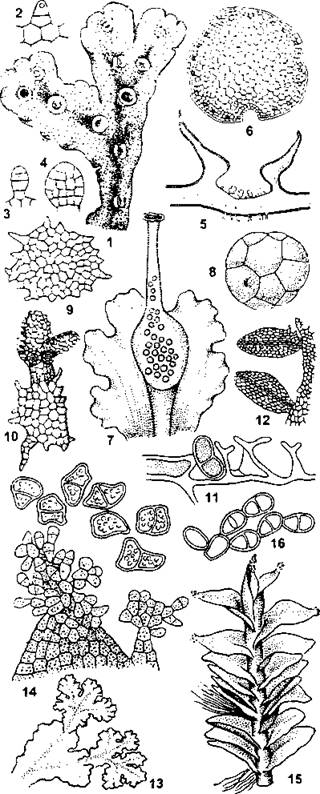
Рис. 198. Различные формы вегетативного размножения у печеночников.
Маршанция (Marchantia polymorphe): 1 - часть слоевища с выводковыми корзиночками; 2-4 - стадии развития выводкового тельца; 5 - выводковая корзиночка в разрезе; 6 — зрелое выводковое тельце. Блазия (Blasia pusilla): 7 - часть слоевища с выводковой колбочкой; 8 - выводковое тельце из колбочки; 9 - звездчатое выводковое тельце с поверхности слоевища; 10 - прорастающее звездчатое выводковое тельце. Рикардия (Riccardia): 11 - эндогенные выводковые тельца. Метцгерия (Metzgeria): 12- выводковые побеги. Пеллия (Реlliа): 13 - выводковые побеги. Сфенолобус (Sphenolobus): 14 - выводковые тельца на листьях. Милия (Mylia anomale): 15- выводковые тельца на листьях; 16- зрелые выводковые тельца (по И. И. Абрамову и соавт.)

Рис. 199. Вегетативное размножение листостебельных мхов. Трахицистис (Trachycystis flagellaris):
1 - растение с выводковыми ветвями. Дикранум (Dicranum flagellare): 2 - выводковая веточка. Плагиотециум (Plagiothecium elegans): 3 - выводковая веточка. Платигириум (Platygyrium repens): 4 - прорастающая и опавшая выводковые почки. Полия (Pohlia annotina): 5 - прорастающая выводковая почка. Дикранум (Dicranum tauricum): б - ломкий лист, образующий протонему и выводковое тело. Тортула (Tortula pagorum): 7 - выводковые листочки на верхушке стебля.Гиофила (Hyophila involuta): 8 - выводковое тело, отделяющееся от подставки. Тетрафис (Tetraphis pellucida): 9 - растения спороносящее, стерильное и с выводковой корзиночкой; 10- выводковое тело. Полия (Pohlia proligera): 11 - выводковые почки. Плагиотециелла (Plagiotheciella latebricola): 12 - верхушка листа с выводковыми телами. Гоиммия (Grimmia anomala): 13 - ранняя стадия образования выводковых тел на верхушке листа. Диходонциум (Dichodontium pellucidum): 14 - выводковое тело. Дикрановейсия (Dicranoweisia cirrata): 15 - прорастающее выводковое тело. Гаплогимениум (Haplohymenium triste): 16- ломкий лист; 17 - прорастающая верхушка листа (по И. И. Абрамову и соавт.)
Происхождение мохообразных до сих пор остается неясным. Хороню сохранившиеся остатки обнаруживаются в отложениях каменноугольного периода, но высокая развитость ископаемых остатков указывает на более ранее появление их предковых форм (по-видимому, в середине палеозойской эры). Непосредственный предшественник мохообразных также не определен, и возможные кандидаты вызывают активные дискуссии среди специалистов-бриологов. По мнению части из них, выраженная индивидуальность мохообразных (их морфологическое строение, преобладание гаметофитного поколения, сильная редукция спорофита и др.) говорит об индивидуальном происхождении этих растений от не установленной формы водорослей.
Согласно другой гипотезе (более распространенной) мохообразные имеют общих предков с другими высшими растениями. На это указывает ряд общих черт, которые, несмотря на очевидные различия, все же имеются. В частности, в спорофитах многих мохообразных обнаружены настоящие (хоть и не функционирующие) устьица, строение которых гомологично устьицам других высших растений. Этот факт позволяет допустить наличие более или менее развитой проводящей системы у предков мохообразных, поскольку трудно понять наличие устьиц без их тесной связи с проводящими структурами. Другим аргументом в пользу общего происхождения всех высших растений, включая мохообразных, является принципиально сходное устройство органов размножения (как полового, так и бесполого). В качестве наиболее вероятных предков сторонники этой гипотезы называют древние примитивные высшие растения - риниофиты, одна из ветвей которых, обитая в условиях обильного увлажнения, утратила проводящую систему. Одновременно с этим происходило упрощение спорофита и, напротив, усложнение гаметофита.
Значение мохообразных в природе довольно ограничено и, в основном, заключается в почвообразовании. Сами эти растения не испытывают потребности в питательном субстрате в той степени, как это присуще другим высшим растениям, поскольку они не обладают корнями и необходимые им вещества способны всасывать всей поверхностью тела. Поэтому мохообразные одними из первых (наряду с лишайниками) заселяют голые скалы и другие места, доселе лишенные растительности, постепенно покрывая их сплошным ковром. Мохообразные мало уязвимы от биотических факторов, потому что очень слабо поражаются микроорганизмами, крайне неохотно поедаются насекомыми, птицами и травоядными животными (чаще всего они их просто не трогают вовсе) и, несмотря на медленный рост (который исчисляется всего лишь несколькими миллиметрами в год), в итоге дали колоссальные залежи торфа, который образуется из-за неполного гниения постепенно отмирающих нижних частей растений (рис. 200). Кроме того, мохообразные участвуют в регуляции водного баланса, так как они задерживают воду и переводят ее в состояние грунтовой. Нередко это приводит к заболачиванию почв и соответственно ухудшению их продуктивности. Следует отметить также способность мохообразных накапливать в своем теле соли тяжелых металлов и радионуклеиды.

Рис. 200. Сфагновые мхи:
1 - сфагнум магелланский (Sphagnum magellanicum); 2 - сфагнум бурый (Sphagnum fuscum); 3 - сфагнум ленский (Sphagnum lenense); 4 - сфагнум Онгстрёма (Sphagnum aongstroemii); 5 - сфагнум балтийский (Sphagnum balticum): 6 - сфагнум дубравный (Sphagnum nemoreum); 7 - сфагнум оттопыренный (Sphagnum squamosum); 8 - сфагнум береговой (Sphagnum riparium); 9 - сфагнум прорезной (Sphagnum perfoliatwn) (по «Жизнь растений», т. 4)
Использование мохообразных в хозяйственной деятельности человека более значительно и, в основном, основывается на эксплуатации торфа. По залежам торфа Россия занимает первое место в мире. На отдельных месторождениях толщина торфяного пласта может быть весьма значительной и достигать нескольких метров (до десяти). Чаще всего торф используют в сельском хозяйстве в качестве органического удобрения и как мульчирующий субстрат для улучшения аэрационного качества тяжелых глинистых почв, делая их структуру более «легкой». Торф используется как топливо. Кроме того, торф активно применяется в строительной индустрии благодаря своим высоким теплоизолирующим свойствам. Менее широко мохообразные используются в медицине. В частности, сфагнум обладает бактерицидным свойством и хорошо впитывает, что позволило широко применять его во время Второй мировой войны (дешевизна и распространенность мха была в тот период особенно важна в условиях повсеместного дефицита медикаментов).
Вопросы для самоконтроля и повторения
1. Какое поколение преобладает в жизненном цикле мохообразных?
2. Как организованы их гаметофит и спорофит?
3. Чем отличается строение гаметофитов печеночных и листостебельных мхов?
4. Как осуществляется половое размножение мохообразных? Какое поколение размножается половым способом?
5. Какие половые органы развиваются у мохообразных?
6. Как происходит оплодотворение у мохообразных?
7. Опишите строение спорофита мохообразных. Откуда спорофит получает питательные вещества?
8. Каким способом размножается спорофит?
9. Как происходит развитие и распространение спор?
10. Как прорастает спора и развивается гаметофит?
11. Как осуществляется вегетативное размножение мохообразных?
12. Какова роль мхов в образовании торфа?