Этот отдел включает в себя группу растений, у которых преобладающим поколением является спорофит. Морфологическое строение папоротникообразных значительно более сложно, чем мохообразных, однако это относится исключительно к спорофиту. Гаметофит, напротив, в значительной мере упростился.
Спорофиты всех современных папоротникообразных имеют прекрасно развитую проводящую систему. Полагают, что наличие этой системы и предопределило гегемонию диплоидного поколения. В свою очередь, появление и развитие проводящих элементов напрямую связано с расчленением тела папоротникообразных на вегетативные органы, главными из которых являются корень и побег (подробнее об органах растений рассказано в соответствующем разделе), и направленный транспорт веществ из одного органа в другой требует наличия соответствующих специализированных структур, соединяющих эти органы. Все органы папоротникообразных являются истинными, поскольку развиваются на спорофите (в отличие от вегетативных органов мохообразных, принадлежащих гаметофиту).
Папоротникообразные являются первыми сосудистыми растениями на Земле. Однако проводящие элементы ксилемы у них за редким исключением представлены не сосудами, а трахеидами. Флоэма также устроена довольно просто и состоит из ситовидных клеток, у которых ситовидные поля разбросаны по всей поверхности клеточной стенки. По уровню развития проводящих элементов данный отдел весьма геторегенен и, наряду с откровенно примитивными вариантами, можно обнаружить формы, у которых проводящие структуры достигли серьезного совершенства. Как правило, они образуют внутри органов четко выраженные проводящие пучки. В подавляющем большинстве случаев между флоэмой и ксилемой в пучках отсутствует камбий, и растения не способны к вторичному утолщению. Не исключено, что отсутствие камбия носит вторичный характер, поскольку у многих ископаемых древовидных папоротникообразных вторичное утолщение имело место. Наличие организованной проводящей системы привело к появлению осевого цилиндра - стели. Причем у разных папоротникообразных можно обнаружить различные типы осевых цилиндров - от примитивных (протостель) до весьма сложно устроенных.
Споры развиваются в спорангиях на листьях (спорофиллах) из поверхностных клеток - эпидермальных и субэпидермальных. Такие листья у одних форм могут одновременно фотосинтезировать, а у других специализируются и морфологически отличаются от трофофиллов. В последнем случае спорофиллы обычно собираются в группы - колоски, или стробилы, занимающие терминальное положение на побеге. Внутри спорангия из спорогенной ткани развиваются споры. Необходимые для своего развития вещества они получают из выстилающего спорангий слоя клеток тапетума. Сам спорангий защищен стенкой, которая предохраняет от высыхания спорогенную ткань.
Папоротникообразные подразделяются на равноспоровые и разноспоровые, в зависимости от соотношения размеров формирующихся спор. У равноспоровых все споры одинаковые и прорастающий из них гаметофит является однодомным. Такие споры образуются в спорангии из материнских клеток спор, которые, делясь мейозом, дают начало тетраде (четырем) совершенно одинаковых спор. У разноспоровых формируются спорангии двух типов - микроспорангии и мегаспорангии. В микроспорангиях из материнских клеток спор в больших количествах образуются тетрады мелких спор - микроспоры, из которых впоследствии прорастают мужские гаметофиты с антеридиями. В мегаспорангиях, также в результате мейоза, из материнских клеток спор образуются очень крупные мегаспоры. Их всегда немного (часто только одна), потому что большая часть спор погибает. Уменьшение количественного выхода мегаспор, очевидно, вызвано тем обстоятельством, что на их образование и развитие затрачивается значительно больше органического материала, нежели на аналогичные процессы формирования микроспор. Согласно эволюционной теории от разноспоровых папоротникообразных произошли все семенные растения.
Сформировавшаяся зрелая спора защищена оболочками. Обычно их две: внутренняя - эндоспорий и наружная - экзоспорий. Причем поверхностный рисунок экзоспория имеет важное систематическое значение, поскольку разнится у представителей разных видов.
Если спорофит папоротникообразных развит очень хорошо и имеет весьма сложное внешнее и внутреннее строение, то гаметофит, напротив, в значительной мере редуцирован. Чаще всего он имеет вид нежной зеленой пластинки или нити и называется заростком. У некоторых форм гаметофит редуцирован еще в большей степени, при этом он не покидает споры, не фотосинтезирует и живет за счет питательных веществ, накопленных в процессе формирования споры. На нижней поверхности заростка образуются нитевидные ризоиды и развиваются немногочисленные половые органы (часто в единичном числе) - архегонии и антеридии. У равноспоровых на одном заростке формируются половые органы обоих типов (однодомный гаметофит), у разноспоровых из микроспоры прорастает мужской гаметофит, на котором развиваются антеридии, а из мегаспоры прорастает женский гаметофит, дающий начало архегониям. Образующиеся в больших количествах дву- или многожгутиковые сперматозоиды выходят из антеридия. Затем сперматозоид проникает в архегоний и сливается там с яйцеклеткой, в результате чего образуется диплоидная зигота. Оплодотворение у папоротникообразных возможно лишь при наличии капельно-жидкой воды, что также имеет аналогию с мохообразными. Именно по этой причине большинство папоротникообразных тяготеют к местам с высокой степенью влажности, хотя и в меньшей степени, чем мохообразные.
Зигота многократно делится, и в итоге образуется многоклеточный зародыш, состоящий из зародышевых органов: корешка, стебелька и листочка, кроме того, зародыш имеет ножку, проникающую в ткань гаметофита. Через ножку неспособный к самостоятельному синтезу органических веществ зародыш получает необходимые для своего развития соединения из фотосинтезирующего гаметофита. У большинства папоротникообразных гаметофит живет недолго, и как только на развивающемся спорофите появляются функционирующие корни и листья, постепенно отмирает (обычно на это уходит несколько недель, исключение составляют плауны, у которых гаметофит живет до нескольких лет).
Современные папоротникообразные в большинстве являются травянистыми растениями, хотя в тропических лесах произрастают и древовидные формы. Однако в предыдущих геологических эрах древовидные папоротникообразные составляли основу лесов. В условиях теплого и очень влажного климата они заселяли многочисленные болота, и конкуренция за наиболее освещенные места привела к возникновению массивных растений высотой до 30 м и нескольких метров в поперечнике. Разумеется, это оказалось возможным благодаря наличию в проводящих пучках камбия, обеспечивающего вторичный рост. Появление древовидных форм отмечено в отложениях девонского периода палеозойской эры. В последующем за ним каменноугольном периоде древовидные папоротникообразные доминировали среди живших в ту эпоху растительных организмов и их остатки со временем сформировали залежи каменного угля, которые в некоторых местах достигают колоссальной толщины.
Несмотря на то что значительная часть папоротникообразных вымерла в предыдущих геологических эпохах, не выдержав конкуренции с более высокоорганизованными семенными растениями, до нашего времени благополучно дожили разнообразные представители трех основных классов этого отдела: папоротниковидные, клинолистовидные (членистые) и плауновидные. Кроме них, в настоящее время на Земле живут еще четыре вида из класса псилотовых, имеющих очень простую организацию.
Тело папоротниковидных расчленено на корень и побег. Следует отметить, что корень имеется не у всех представителей класса. В частности, он отсутствует у сальвиний, но это носит вторичный характер и связано со средой обитания. Зародышевый корешок рано отмирает, и корни папоротниковидных отрастают на стебле и листьях, т.е. по своему происхождению они являются придаточными. Нередко корни ветвятся и в итоге образуют корневую систему, достаточно мощную, чтобы надежно снабжать водой и минеральными веществами надземную часть растения.

Рис. 201. Регнеллидиум двулистный (Regnellidium diphyllum) (по М.Д. Махлину и соавт.)

Рис. 202. Лигодиум вьющийся (Lygodium volubile), часть листа (по А. Е. Боброву)
Стебли папоротниковидных, за исключением древовидных форм, развиты относительно слабо и часто почти полностью располагаются в почве. Морфологически они довольно разнообразны и могут быть ползучими (рис. 201) или вьющимися (рис. 202) (такие стебли еще называют корневищами) или прямостоячими (у древовидных форм) (рис. 203). Длина стебля у разных форм варьирует от нескольких миллиметров до 25 м. Укороченные стебли некоторых папоротников напоминают клубни (рис. 204). У многих папоротников стебель ветвится.
Проводящие элементы стебля папоротниковидных образуют хорошо развитые осевые цилиндры - стелы (см. рис. 117). У большинства ныне живущих папоротников имеются различные типы сифоностелы (рис. 205), но у отдельных форм можно встретить и другие стелы (от примитивной протостелы до весьма совершенной эустелы). Наличие хорошо развитых листьев у папоротниковидных объясняет присутствие обширных листовых следов в стеле. Напоминаем, что лакуной, или листовым прорывом, называется участок осевого цилиндра, который располагается над областью отхождения листового следа и заполнен паренхимой.


Рис. 203. Древовидный папоротник циатея грязноватая (Cyathea contaminans) в горном лесу на острове Ява (по В. Н. Гладковой)

Рис. 205. Поперечный срез корневища матонии гребневидной (Matonia pectinata); видна полициклическая сифоностела (по В. Н. Гладковой)
На листья у папоротниковидных приходится основная часть биомассы растения. Как правило, они имеют очень крупные размеры, т. е. для папоротниковидных характерна макрофиллия (у некоторых видов длина листьев может превышать 30 м). Раньше (а нередко и в настоящее время) большие листья папоротников называли вайями (греч. baion - пальмовая ветвь). Полагают, что они возникли в результате дорзо-вентрального уплощения целых ветвей предковых форм (вероятнее всего риниофитов), доказательством чему служат ископаемые остатки древних папоротников, у которых наблюдаются переходные стадии уплощения целых крупных ветвей. Кроме того, несомненным аргументом в пользу сказанного служит наличие апикальной меристемы у листьев, что обеспечивает им (подобно стеблям) верхушечный рост.
Морфология листа у различных папоротниковидных весьма разнообразна. Прежде всего это относится к форме и размерам. За исключением относительно немногих форм с сидячими листьями, у большинства папоротников листья имеют выраженный черешок, посредством которого они соединяются со стеблем. Очень редко листовая пластинка остается цельной, у подавляющего большинства представителей класса она более или менее сложно расчленена. В наиболее примитивном варианте оно дихотомическое (что неудивительно, если учесть происхождение самого органа от уплощенных ветвей предшественников, которые сами ветвились сходным образом), но значительно чаще листья имеют одно-, дву- или многоперистое расчленение. При этом стержень листа, являющийся продолжением черешка, называется рахисом, а остальные оси, находящиеся на нем (если лист дву- или многоперистый), - черешочками (рис. 206). Сами листовые пластинки (они называются перьями – пластинки от первой до последней генерации или перышками — пластинки последней генерации) могут быть цельными или в определенной мере рассеченными.
Внутри листа обнаруживается хорошо развитая проводящая система, образованная проводящими пучками. Проекция пучков на поверхности носит название жилок листа, а их совокупность называется жилкованием. У примитивных форм оно дихотомическое и не образует внутри листовой пластинки сплошной сети (специалисты называют такое жилкование открытым). Поэтому появившееся значительно позже сетчатое жилкование считается более прогрессивным, поскольку формирует внутри листа густую сеть из проводящих пучков с многочисленными перемычками между ними. Такой тип жилкования позволяет эффективнее транспортировать необходимые для жизнедеятельности растения вещества.

Рис. 206. Строение листа папоротника (схема):
1 - черешок: 2 - пластинка листа; 3 - перо первого порядка; 4 - перышки (или перья второго порядка); 5 - лопасть перышка; 6 - рахис (по А. Л. Тахтаджяну)

Рис. 207. Оноклея (Опосієа strutbiopteris): 1 - спорофилл; 2 - трофофилп (по Н.А.Комарницкому и соавт.)
Листья большинства папоротниковидных совмещают две функции: фотосинтетическую и репродуктивную. Однако у отдельных форм встречается специализация. В этом случае часть листьев служит для развития спорангиев, утрачивая при этом хлорофилл и способность к фотосинтезу. Такие листья называются фертильными (лат. fertilis - плодородный, плодоносный, плодовитый), или спорофиллами (греч. spora - семя, phyllon - лист). На других листьях - трофофиллах (греч. trophe - питание, phyllon - лист) - спорангии не образуются, и они, активно фотосинтезируя (благодаря наличию хорошо развитой хлоренхимы), снабжают органическими соединениями все тело растения, в том числе и фертильные листья со спорангиями (рис. 207).
Спорангии на листьях располагаются тремя основными способами. У наиболее примитивных ископаемых форм органы бесполого размножения занимают верхушечное расположение на листьях, подобно спорангиям ринио- фитов (разумеется, листья у последних еще отсутствовали, и данная аналогия указывает на терминальное расположение спорангиев на теломах предков папоротниковидных). Среди современных папоротников отсутствует строго терминальное расположение спорангиев, но у самых простых (ужовниковые) можно встретить близкие к этому варианты (рис. 208).

Рис. 208. Гельминтостахис (Helmintostachys zeylanica) (по Н. А. Комарницкому и совет.)
Более поздним стало краевое расположение спорангиев на листовой пластинке (рис. 209). Специалисты связывают этот тип расположения с уплощением ветвей ри- ниофитов в процессе эволюции листа, в результате чего спорангии оказывались оттесненными на край органа.

Рис. 209. Схизейные и гименофилловые:
I - схизея: А - общий вид Schizaea pusilla; В - стадии развития спорангиев (по краю листа Sen. rupestris); В - спорангий Schizaea bifida (кольцо верхушечное, с продольной трещиной); II - гименофипл (Hymenophyllum dilatatum): А - общий вид (1 - расположенные по краю листа спорангии); Б - спорангии с полным косым кольцом; III - трихоманес (Thchomanes reniforme) (по Н. А. Комарницкому и соавт.)
Среди современных папоротников краевое нахождение спорангиев распространено довольно широко.
Самым прогрессивным способом расположения, несомненно, является поверхностное, которое появилось позднее всех других и у современных папоротников наиболее распространено. В этом случае спорангии развиваются не на верхушках листьев или их краях, а на нижней поверхности листа (рис. 210). Такое расположение в наименьшей степени мешает фотосинтезу (поскольку он наиболее интенсивен на верхней, наиболее освещенной стороне листа) и в наибольшей защищает спорангий.

Рис. 210. Общий вид и расположение сорусов на листьях папоротников семейства граммитисовых:
1 - граммитис усыпанный (Grammltis adspersa) - общий вид; 2 - граммитис усыпанный - часть листа с сорусами (увел.); 3 - ктеноптерис небольшой (Ctenopteris subminuta) - перо (увел.); 4 - ксифоптерис Иеронима (Xiphopteris hieronimiisii) - часть листа с сорусами (увел.); 5 - ксифоптерис рассеянноволосистый (Xiphopteris sparsipiiosa) - часть листа с сорусами (увел.) (по Т. Д. Суровой)
Закладка спорангия начинается с обособления от общей массы особых инициальных клеток, из которых в последующем разовьется спорангий (рис. 211).

Рис. 211. Осмунда (Osmunda regalis):
1 - разделение листьев на бесплодные и спороносные сегменты; 2-5 - стадии развития спорангия; 6 - зрелый вскрывшийся спорангий (no Н. А. Комарницкому и соавт.)
В более примитивном случае инициальных клеток много и спорогенная ткань оказывается окруженной многоклеточной оболочкой. Среди ныне живущих папоротников такое встречается нечасто. У большинства современных форм начало будущему спорангию дает лишь одна инициальная клетка. В результате спорангий оказывается окруженным однослойной стенкой (см. далее рис. 213-П). Такой способ считается более продвинутым, потому что большая часть усилий растения затрачивается не на формирование стерильной стенки, а непосредственно на развитие спорогенной ткани.
У просто организованных папоротников (формы с терминальным и краевым типами расположения) спорангии располагаются поодиночке. И лишь у видов с поверхностным расположением спорангия последние закладываются группами - сорусами (греч. soros - куча) (рис. 212). В большинстве случаев спорангии в сорусе защищены выростом листа - покрывалом, или индузием (лат. indusium — верхняя туника), и лишь наиболее просто устроенные скопления спорангиев не имеют защитных структур. Развитие спорангиев в сорусе может быть одновременным или осуществляться в разные сроки. Полагают, что в последнем случае растения имеют некоторые преимущества. В частности, одновременное созревание спор может совпасть с ухудшением условий обитания растения, что может быть причиной гибели всего генеративного материала. Что касается последовательного развития спорангиев, то здесь выход спор происходит по мере их созревания, и процесс оказывается более или менее пролонгированным. При изменении действия различных факторов окружающей среды это дает шанс хотя бы для части спор оказаться в выгодных для их прорастания условиях.

Рис. 212. Мужской папоротник (Dryopteris filix-mas):
I - взрослое растение (спорофит) с молодыми улиткообразно скрученными листьями; II - часть листа с сорусами, одетыми покрывалами (1 - сорус); III - поперечный разрез через лист с сорусом (2 - индузий, 3 - спорангии); IV - спорангий с кольцом (4 - кольцо) (по Н.А. Комарницкому и соавт.)
Выход спор из созревших спорангиев у наиболее примитивных папоротников осуществляется через округлое или щелевидное отверстие, расположенное на верхней части спорангия (см. рис. 211-6). Среди современных папоротниковидных это встречается редко. У большинства же на стенке спорангия образуются специализированные структуры, обеспечивающие его раскрытие. Они представляют собой группу клеток с неравномерно утолщенными стенками и в своей совокупности называются кольцом (рис. 213). У разных папоротников кольца могут иметь самые разнообразные особенности, при этом выделяют три основных типа колец - поперечное, косое и продольное.

Рис. 213. Лептоспорангиатные папоротники:
I - спорангий с кольцом и раскрывание спорангия: А - зрелый спорангий; В - раскрывшийся спорангий; В - клетки кольца; II - развитие спорангия (1 - клетка, дающая начало спорангию; 2 - клетка, образующая ножку спорангия; 3 - выстилающий слой; 4 - археспорий) (по Н. А. Комарницкому и соавт.)
Большая часть папоротников продуцируют одинаковые по размеру и форме споры, т. е. являются равноспоровыми растениями. Однако имеется немало разноспоровых папоротников, например сальвиниевые. Полагают, что разноспоровость возникала несколько раз у различных групп ископаемых папоротниковидных.
Строение споры достаточно типично, хотя и чрезвычайно разнообразно в деталях. Покровы образуют две оболочки: внутренняя - интина (лат. intus - внутри) и наружная - экзина (англ, exine extine, лат. extimus - внешняя оболочка). Последняя как раз и демонстрирует широкую морфологическую вариабельность у разных групп. Это обстоятельство охотно используют систематики для решения спорных вопросов, касающихся таксономического положения сходных по своему строению форм различных папоротников.
Споры рассеиваются в очень больших количествах, но громадное большинство из них попадает в агрессивную среду и погибает. Лишь небольшая часть спор оказывается в благоприятных условиях, и после определенного периода покоя они прорастают. Следует отметить, что не всегда перед прорастанием споры выдерживают паузу. У ряда форм, обитающих во влажном климате в дождевых тропических лесах, споры имеют хлоропласты и поэтому окрашены в зеленый цвет, хотя у большинства папоротников споры имеют темную окраску, потому что содержат не хлоропласты, а их предшественники - пропластиды. Такие споры прорастают сразу. Иногда зеленые споры продуцируют папоротники, произрастающие в менее влажном климате, в частности умеренном, но у них это происходит не регулярно, а лишь в случае затяжных дождей.
Споры прорастают при высокой влажности и температуре выше +15°С. Кроме того, споры большинства папоротников для прорастания нуждаются в свете. В ряде случаев прорастание осуществляется как на свету, так и в темноте или только в темноте.
Из споры прорастает гаметофит (рис. 214). У равноспоровых папоротников он представляет собой вполне самостоятельное растение с хорошо развитой ассимилирующей тканью, а потому способен сам снабжать себя необходимыми органическими соединениями. Однако даже такой гаметофит очень нежен и лишен покровов, защищающих его от высыхания, по этой причине гаплоидная фаза папоротников развивается только в условиях достаточного увлажнения. Морфологически гаметофит большинства равноспоровых папоротников имеет вид зеленой пластинки, уплощенной в дорзо-вентральном направлении. Очень часто он имеет сердцевидную форму (между лопастями такого гаметофита находится точка роста) (см. далее рис. 216-А), хотя это имеет место далеко не у всех папоротников (рис. 215, см. рис. 214). Обычно гаметофит очень маленький - всего около 0,5 см в поперечнике, но у наиболее примитивных представителей гаметофит может достигать относительно значительных размеров (до 5 см) и иметь разнообразную форму, к тому же он гораздо более долговечен. На вентральной (обращенной к субстрату) поверхности гаметофита образуются многочисленные нитевидные ризоиды, которые связывают его с субстратом, а также способствуют поглощению воды.

Рис. 214. Развитие гаметофита давалаии мелкозубчатой (Davallia denticulata):
1, 2, 3 - стадии развития нитчатой формы гаметофита; 4, 5- формирование лопатчатого гаметофита; 6 - зрелый гаметофит с несколькими архегониями и антеридиями; 7, 8 - стадии развития архегония; 9, 10, 11, 12, 13 - стадии развития антеридия; 14 - молодой спорофит на гаметофите (no Т. Д. Суровой)

Рис. 215. Гаметофиты схизейных:
1 - актиностахис малокопосый (Actinostachys oligostachys); 2 - схизея маленькая (Schizaea pusilla); 3 - мория кафрская (Mohria caffrorum); 4 - лигодиум японский (Lygodium japonicum) (по T. Н. Поповой)
На гаметофите равноспоровых папоротников развиваются половые органы (рис. 216-А). Как правило, на одном растении имеются как антеридии, так и архегонии, т.е. гаметофит однодомен. При этом у большинства папоротников мужские органы возникают несколько раньше женских. Как показали исследования, этим управляют особые соединения, обладающие активностью гормонов. Более раннее развитие антеридиев называется протандрией, в отличие от протогинии, когда раньше развиваются архегонии.

Рис. 216. Заросток и молодой спорофит папоротников:
А - мужской папоротник (Dryoptehs filix-mas); 1 - заросток; 2 - развитие архегония; 3 - зрелый антеридий; 4 — антеридий в момент выхода многожгутиковых сперматозоидов; 5 - сперматозоид; 6 - Adianthum (поперечный разрез заростка и зародыша): 6 - заросток; 7 – ризоиды; 8 - неоплодотворенные архегонии; 9 - первый лист зародыша - «семядоля»; 10 - первичный корень; 11 - ножка (по Н. А. Комарницкому и соавт.)
Количество половых органов, образующихся на одном гаметофите папоротниковидных, относительно небольшое.
При этом у более высокоорганизованных форм отмечается определенное упрощение их строения. В особенности это относится к мужским органам. В частности, у примитивных папоротников в крупных антеридиях, погруженных в ткань гаметофита, образуются многочисленные сперматозоиды. У более сложно организованных форм антеридии возвышаются на подставках и имеют меньшие размеры, соответственно количество развивающихся в них сперматозоидов значительно меньше.
Как и у всех папоротникообразных (а также мохообразных), оплодотворение папоротниковидных осуществляется только в присутствии капельножидкой воды. Снабженные многочисленными жгутиками и потому подвижные гаплоидные сперматозоиды выходят из антеридиев и, привлекаемые химическими соединениями, которые выделяют архегонии, подплывают к ним, проникают через шейку в брюшко и сливаются с находящейся там гаплоидной яйцеклеткой. В результате образуется зигота с диплоидным набором хромосом в ядре. Несмотря на количество оплодотворенных яйцеклеток, лишь одна зигота развивается и дает начало многоклеточному зародышу. Остальные при этом погибают и не мешают развитию молодого спорофита, который первоначально нуждается в значительном количестве органического материала для построения своего тела. Такое течение событий представляется весьма рациональным, если учесть, что гаметофит изначально имел небольшие размеры и, безусловно, не смог бы обеспечить развитие сразу нескольких зародышей. Вскоре у растущего зародыша образуются функционирующие корни и фотосинтезирующие органы, после чего он способен самостоятельно жить и становится независимым от материнского гаметофита (рис. 216-Б). Сам гаметофит вскоре после этого отмирает. Схема жизненного цикла равноспорового папоротника показана на рис. 217.

Рис. 217. Схема жизненного цикла равноспорового папоротника Dryopteris filix-mas (no Н. Гоину и соавт., с изменениями)
Из всех папоротникообразных папоротниковидные являются наиболее процветающей группой. Несмотря на то что многие виды ранее вымерли, в настоящее время класс насчитывает не менее 10 000 видов, заселяющих самые разнообразные экологические ниши. Из-за потребности в воде для оплодотворения папоротниковидные, в основном, заселяют затененные влажные места, где могут быть весьма многочисленными. Однако на Земле живут и другие формы папоротников. В частности, актиниоптерис южный является выраженным ксерофитом и заселяет засушливые области Африканского и Азиатского континентов. Среди папоротников есть и водные формы (здесь следует иметь в виду, что водный образ жизни для этих растений представляет собой явление вторичное), например салъвиния и др. (рис. 218).

Рис. 218. Водные папоротники:
I - марсилея четырехлистная (Marsilea quadrifolia) в аквариуме;
II - сапьвиния продолговатолистная (Salvinia oblongifolia);
III - цератоптерис василистниковидный (Ceratopteris thalictroides) в аквариуме; IV - сапьвиния ушковидная (Salvinia auriculata); V- азолла каролинская (Azolla caroliniana) (по «Жизнь растений», т. 4)
Интересную группу составляют эпифитные формы, которые живут на стволах деревьев, используя их в качестве подставки и опоры (рис. 219). Кроме того, среди папоротников есть немало лиан. В тропических лесах произрастают немногочисленные ныне (но очень широко распространенные в предыдущие геологические эпохи) древовидные папоротники.

Рис. 219. Папоротники-эпифиты:
I - даваллия крыночковидная (Davallia pyxidata); II - ужовник повислый (Ophioglossum pendulum); III - асплениум гнездовый, или птичье гнездо (Asplénium nidus); IV - гименофилл Райта (Hymenophyllum wrightii); V - трихоманес маленький (Trichomanes minutum); 6 - леммафиллум мелколистный (Lemmaphyllum microphyllum) (по «Жизнь растений», т. 4)
Тело плауновидных расчленено на побег и корень (рис. 220). Зародышевый корешок рано отмирает, и поэтому корневая система этих растений сформирована корнями, отходящими от подземной части побега - корневища (напомним, что такие корни называют придаточными). Надземная часть побега современных плауновидных остается травянистой, хотя среди предковых форм имелось и немало древовидных. Верхушечный рост обеих частей побега осуществляется благодаря наличию апикальных меристем, которые, раздваиваясь, обеспечивают ветвление. Оно у плауновидных носит дихотомический характер и может быть равным и неравным. Осевые цилиндры могут быть различных типов, и в процессе индивидуального развития происходит смена одного другим. При этом листовые прорывы в осевом цилиндре отсутствуют (рис. 221).

Рис. 220. Плаун булавовидный (Lycopodium clavatum):
А - общий вид;
Б - спорофилл;
В - поперечный разрез стебля;
1 - эпидермис; 2 - кора; 3 - листовой след; 4 - флоэма; 5 - эндодерма; 6 - ксилема; 7 - механическое кольцо (по Н. А. Комарницкому и соавт.)
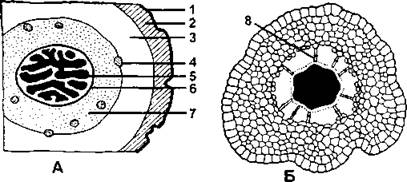
Рис. 221. Поперечные срезы стеблей плауновидных:
А - срез стебля плауна булавовидного (Lycopodium clavatum): 1 - эпидерма; 2 - внешняя кора; 3 - срединная кора; 4 - листовой след; 5 - проводящие элементы ксилемы; 6 - эндодерма; 7 - внутренняя кора; Б - срез стебля селагинеллы селаговидной (Selaginella selaginoides); 8 - трабекула (по В. Р. Филину)
Листья на стебле обычно располагаются по спирали, в отдельных случаях наблюдается супротивное, или мутовчатое, листорасположение. Для плауновидных характерна микрофилия, т. е. листья имеют небольшие размеры с единственной центральной жилкой (см. рис. 126). У некоторых представителей у основания листа имеется вырост в виде пластинки - язычок, или лигула. По своему происхождению листья плауновидных отличаются от листьев всех других высших растений, поскольку представляют собой выросты стебля (энации, поэтому такие листья еще называются энацион- ными). Листья могут быть вегетативными - трофофил- лы или спороносными - спорофиллы (см. рис. 220-Б). Морфологические отличия между обоими типами листьев могут быть существенными или (что встречается реже) отсутствовать вовсе. У примитивных ископаемых плауновидных вегетативные и спороносные листья располагались на стебле без особого порядка (рис. 222), однако у современных форм спорофиллы образуют скопления в виде спороносных зон (если такие листья собраны в средней части стебля) или стробил (если спорофиллы собраны на концевых участках веточек).

Рис. 222. Ископаемые плауновидные:
А - содония украшенная (Sawdonia ornata): 1 - участок побега с шипами и спорангиями; Б — каулангиофитон иглистый (Kaulangiophyton akantha): 2 - участок побега с шипами и спорангиями; В - дрепанофикус колючковидный (Drepanophycus spinifornils) (по Н. С. Снигиревской)
Среди плауновидных имеются как равноспоровые, так и разноспоровые формы. Равноспоровыми являются плауновые. Одиночные спорангии у них обычно развиваются в пазухах спорофиллов (рис. 223), которые часто бывают собраны в стробилы (рис. 224, см. рис. 220). Развитие спорангия и созревание в нем спор продолжается достаточно долго и занимает от нескольких месяцев до нескольких лет. Процесс инициируется группой поверхностных клеток спорофилла. Зрелый спорангий имеет округлую или почковидную форму и посредством короткой ножки связан со спорофиллом. Внутри спорангия находится большое количество материнских клеток спор. В результате деления каждой из них посредством мейоза образуются четыре мелкие споры, ничем между собой не различающихся. Отдельная спора заключена в две оболочки: внутреннюю - интину и наружную - экзину, которая у разных представителей имеет различный рельеф поверхности (рис. 225). В сухую погоду зрелый спорангий раскрывается поперечно ориентированной щелью, которая закрывается при увеличении влажности атмосферного воздуха.

Рис. 223. Схема расположения спорангиев на стебле и щели вскрывания в стенке спорангия у плаунов:
I - плаун запиваемый (Lycopodium inundatum); Il - плаун поникший (Lycopodium cernuum); III - плаун оттопыренный (Lycopodium squamosum); IV - плаун светловатый (Lycopodium lucidulum) (по В. Р. Филину)

Рис. 224. Плауновидные:
I - селагинелла селаговидная (Selaginella selaginoides); II — плаун баранец (Lycopodium selago); III - плаун годичный (Lycopodium annotmum); (по «Жизнь растений», т. 4)

Рис. 225. Споры плаунов и плауновидных под сканирующим электронным микроскопом:
I - споры плауна заливаемого (Lycopodium inundatum) (увел, около 1200):
II - спора плауна булавовидного (Lycopodium clavatum) (увел, около 2000):
III - тетраэдрическая тетрада спор плауна баранца (Lycopodium selago) (увел, около 1200): IV - тетраэдрическая тетрада спор плауна фпегмарии
(Lycopodium phlegmaria) (увел, около 2000); V - мегаспора и тетрады микроспор сепагинеппы селаговидной (Selaginella selaginoides) (увел, около 160):
VI - тетрада микроспор сепагинеппы селаговидной (увел, около 1200) (по «Жизнь растений», т. 4)
Спорангии разноспоровых плауновидных (к ним относятся селагинелловые и полушниковые) не одинаковые (рис. 226). Различают более крупные - мегаспорангии, которые развиваются на мегаспорофиллах, и относительно небольшие - микроспорангии, развивающиеся на микроспорофиллах. Обычно оба эти типа спорофиллов развиваются на одном и том же растении, будучи при этом собранными в стробилы. Как правило, в верхней части стробилы находятся микроспорофиллы, а в нижней - мегаспорофиллы. Значительно реже спорофиллы распределяются беспорядочно.
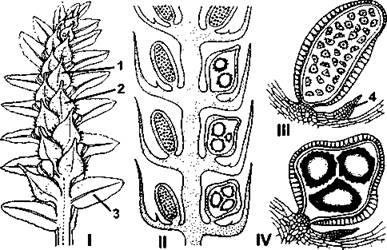
Рис. 226. Строение стробила и спорангиев у селагинеллы:
I - стробил сепагинеппы зопотистостебельной (Selaginella chrysocaulos), вид с нижней стороны: 1 - микроспорофилл, 2 - мегаспорофилл, 3 - боковой лист; II - схема продольного разреза стробила селагинеллы Крауса (Selaginella kraussiana); III - микроспорангий селагинеллы Крауса: 4 - язычок; IV - мегаспорангий селагинеллы Крауса (по В. Р. Филину)
Окруженные различного рода выростами листа спорангии располагаются в его пазухе, где посредством мейоза из материнских клеток образуются споры. Размеры спор изначально неодинаковые и у различных видов колеблются от 20 мкм до 100 мкм у микроспор и от 100 мкм до 1500 мкм у мегаспор. Количество образовавшихся спор может быть различным и широко варьирует у разных форм. Микроспор всегда больше, и в спорангии их может насчитываться до миллиона. Количество мегаспор гораздо меньше и исчисляется десятками или сотнями (у некоторых полушниковых в мегаспорангии могут находиться свыше двух тысяч мегаспор), у некоторых видов в мегаспорангии развивается всего лишь одна мегаспора. Разнообразный рельеф поверхностной оболочки споры служит видовой характеристикой (см. рис. 225).
Выход спор из спорангия осуществляется несколькими способами. В частности, створки спорангия могут традиционно раскрываться, и споры выбрасываются в воздух, увлекаясь при этом ветром, который переносит их на определенное расстояние от материнского растения. У полушников все происходит еще проще - после сезонного отмирания листьев стенка спорангия разлагается и споры оказываются в почве, где распространяются потоками воды или беспозвоночными животными - обитателями почвы (реже более крупными позвоночными). Наконец, у некоторых видов выходят только микроспоры, а мегаспоры вообще не покидают мегаспорангия (подробнее об этом будет рассказано ниже при описании развития гаметофита разноспоровых плауновидных). У многих разноспоровых из спорангиев выходят не споры, а женские и мужские гаметофиты.
Дальнейшее развитие споры у равно- и разноспоровых плауновидных происходит неодинаково. У равноспоровых возможны два варианта прорастания спор. При первом из них (более быстром) спора прорастает уже через несколько дней после очень непродолжительного периода покоя. Молодой гаметофит имеет фотосинтезирующую надземную часть и лишенную хлорофилла подземную, которая вступает в симбиоз с почвенными грибами. Причем, если поблизости отсутствуют гифы гриба, развитие гаметофита прекращается, и он погибает.
У других равноспоровых плауновидных спора находится в состоянии покоя значительно дольше и прорастает через несколько лет. При этом споры постепенно погружаются в субстрат (с током воды, с помощью членистоногих и червей и т. п.) и со временем оказываются на глубине в несколько сантиметров. Из-за отсутствия под землей света гаметофит не может фотосинтезировать и вынужден жить сапрофитно, как и в предыдущем случае, на самых ранних этапах развития, вступая в симбиоз с почвенными грибами, без которых жизнь гаметофита невозможна.
Гаметофит растет очень медленно. Вначале он представляет собой малоупорядоченный комок клеток, но со временем наблюдается организованный рост за счет выделившейся верхушечной меристемы. Появляются ризоиды, в клетки которых проникают гифы гриба. Постепенно гаметофит достигает нескольких миллиметров в поперечнике и приобретает форму, характерную для данного вида (рис. 227). Этот процесс занимает до 15 лет. На верхней стороне гаметофита развиваются половые органы, причем как мужские, так и женские, поскольку заростки равноспоровых плауновидных однодомные. Антеридии и архегонии довольно многочисленны и почти полностью погружены в ткань гаметофита (рис. 228) (над поверхностью возвышаются только шейки архегониев).

Рис. 227. Заростки (гаметофиты), гаметангии, зародыши и проростки плаунов:
А - заросток плауна заливаемого (Lycopodium inundatum);
Б - заросток плауна поникшего (Lycopodium cernuum) с двумя зародышами;
В - заросток и проросток плауна булавовидного (Lycopodium clavatum);
Г - заросток и проросток плауна сплюснутого (Lycopodium compianatum);
Д-Ж- заростки и проросток плауна баранца (Lycopodium selago);
З - заросток плауна фпегмарии (Lycopodium phlegmaria);
И - антеридий плауна булавовидного;
К - архегоний плауна булавовидного; Л - зародыш плауна булавовидного;
М - проросток плауна булавовидного;
Н - схема строения зародыша плаунов, обладающих подземными заростками;
О - схема строения зародыша плаунов, обладающих наземными заростками; 1 - заросток; 2 - корень; 3 - лист; 4 - ножка; 5 - подвесок; 6 - протокорм; 7 - проросток; 8 - протофипл; 9 - яйцеклетка (по В. Р. Филину)
Оплодотворение, как и у всех папоротникообразных, возможно лишь в присутствии капельно-жидкой воды. Антеридии развиваются несколько раньше архегониев, и из них выходят многочисленные сперматозоиды, подвижность которых определяется наличием на переднем конце клетки двух жгутиков (редко жгутиков бывает три). Активно перемещаясь в воде по направлению к архегонию, гаплоидные сперматозоиды достигают последнего, через шейку проникают в брюшко архегония, где один из них сливается с гаплоидной яйцеклеткой. В результате образуется диплоидная зигота.

Рис. 228. Развитие плауна:
А — спора;
Б - подземный заросток: 1 - архегоний; 2 - антеридий; 3 - зародыш; 4 - эпидермис; 5 - гифы гриба; 6 - ризоиды;
В - антеридий и архегоний: I - антеридий; II - сперматозоиды; III - молодой архегоний; IV - вскрывшийся архегоний; 7 - яйцеклетка в брюшке архегония;
Г- строение и развитие зародыша: V- оплодотворенная яйцеклетка (зигота) поделилась поперечной перегородкой, образовав две клетки - 8 и 9; VI - клетка 8 больше не делится и образует подвесок, клетка 9 делится продольной перегородкой; VII - дальнейшее деление клеток зародыша; VIII - зародыш с дифференцированными органами; 8 - подвесок; 9 - зародыш; 10- заросток (гаметофит); 11 - точка роста (апикальная меристема); 12- примордий (зачаток) первого листа; 13 - корень; 14 - ножка (по Н. А, Комарницкому и соавт., с изменениями и дополнениями)
Без выраженного периода покоя зигота приступает к делению, причем оно совершается перпендикулярно вертикальной оси. После этого верхняя клетка уже не делится и трансформируется в подвесок или ножку. Подвесок проникает в ткань гаметофита (т. е. является присоской - гаусторией, лат. haustor - черпающий, пьющий, глотающий) и обеспечивает снабжение нужными веществами зародыш, который в свою очередь развивается из нижней клетки в результате ее многократных делений. У зародыша последовательно образуется стебелек, первый лист (не имеющий проводящего пучка), а в основании листа - корень, который является придаточным, так как образуется не на корне, а на листе.
Наземная часть молодого спорофита зеленеет, и постепенно в ней накапливается значительное количество фотосинтезирующей ткани, достаточное для самостоятельного существования. У форм с наземным гаметофитом эти события происходят в течение одного сезона, а у форм, имеющих полностью сапрофитный подземный гаметофит, процесс может растянуться на несколько лет, поскольку у них побег молодого спорофита преодолевает до поверхности почвы более или менее значительные расстояния. Все это время проросток растет очень медленно, а необходимые для жизни вещества он получает из гаметофита (который при этом также продолжает свой рост) через подвесок.
Прорастание мега- и микроспор происходит несколько иначе. При этом следует отметить, что среди всех, папоротникообразных гаметофит разноспоровых плауновидных наиболее значительно упрощен. В особенности это относится к мужскому гаметофиту, который вообще не покидает споры. Так, у селягинелловых деление ядра микроспоры дает начало двум клеткам: небольшой - проталлиальной (вегетативной, или ризоидальной) и более крупной - антеридиальной, из которой развивается единственный антеридий, к тому же сильно редуцированный. В типичном случае он состоит из однослойной стенки, состоящей из восьми стерильных клеток, которая отграничивает и в определенной мере защищает две или четыре сперматогенные клетки. К этому моменту приурочен выход микроспор из микроспорангия, что осуществляется посредством быстрого образования продольной щели на стенке спорангия, из которой микроспоры энергично выбрасываются наружу на расстояние в несколько сантиметров. В итоге они попадают на землю, где и будут разворачиваться последующие события развития мужского гаметофита. Вначале из немногочисленных сперматогенных клеток путем митотических делений образуются 256 материнских клеток сперматозоидов - сперматидиев. Такой антеридий является зрелым, а все его стерильные клетки ослизняются. В образовавшейся слизи плавают подвижные сперматидии до тех пор, пока оболочка споры не лопается и сперматидии не выйдут наружу. Там из них выходит по одному сперматозоиду, снабженному двумя жгутиками, с помощью которых сперматозоиды в капельно-жидкой воде подплывают к архегонию, проникают через шейку в его брюшко и один из них сливается с находящейся там яйцеклеткой.
У полушниковых мужской гаметофит редуцирован в еще большей степени, чем у селягинелловых. Он по- прежнему состоит из одной проталлиальной клетки и антеридия. Однако стенка антеридия образована не восемью, а всего лишь четырьмя клетками, которые защищают четыре сперматидия. Из них впоследствии выходят четыре сперматозоида, причем их подвижность обеспечивается не двумя жгутиками (как у селягинелл), а многими.
Развитие женского гаметофита происходит внутри мегаспоры. Этому способствует наличие в ней значительных запасов питательных веществ, за счет которых гаметофит растет и развивается. Чаще всего мегаспоры покидают мегаспорангий, но у многих видов развитие гаметофита возможно при нахождении мегаспоры в пределах спорангия (при этом стробилы с мегаспорангиями обычно сами опадают со спорофита на землю, однако у ряда видов развитие гаметофита и даже последующее оплодотворение яйцеклетки в архегоний осуществляется внутри мегаспорангия, находящегося на функционирующем материнском растении). Развитие женского гаметофита разноспоровых плауновидных начинается с многократных митотических делений сначала ядра самой споры, а затем образовавшихся в результате этого дочерних ядер. При этом образуется многоядерная масса, которая лишь впоследствии разделяется межклеточными перегородками, давая начало многоклеточному женскому заростку. Клетки, расположенные в верхней части заростка, делятся более энергично, в результате чего оболочка мегаспоры в этом месте лопается и через образовавшуюся щель женский гаметофит частично вываливается из мегаспоры наружу. Будучи освещенными, клетки заростка зеленеют, и поэтому гаметофит способен самостоятельно снабжать себя органическими соединениями. На незеленой части заростка образуются ризоиды и немногочисленные архегоний с короткими шейками (1 — 3 канальцевые клетки), погруженными в ткань гаметофита.
Развитие зародыша из зиготы у разноспоровых плауновидных широко варьируется. Сам зародыш, будучи полностью сформированным, состоит из стебелька с верхушечной точкой роста, двух листочков, корешка и ножки, с помощью которой зародыш получает из гаметофита необходимые ему вещества.
Зависимость молодого спорофита от питающего его заростка может быть довольно длительной, но в конечном счете корешок и листочек прорывают ткань гаметофита, выходят наружу и, достигнув достаточного уровня развития, самостоятельно снабжают спорофит водой с минеральными веществами и продуктами фотосинтеза. После этого заросток постепенно отмирает. Схема развития разноспоровых плаунов представлена на рис. 229.
Современных плауновидных насчитывается значительно меньше, чем папоротниковидных, но в предыдущие геологические эпохи (особенно в каменноугольном периоде палеозойской эры) они были весьма широко представлены в фитоценозах (растительных сообществах) того времени, образуя заросли и даже леса. В начале мезозойской эры подавляющее большинство плауновидных (и практически все крупные формы) вымерли, не выдержав конкуренции с более прогрессивно организованными голосеменными растениями. Среди ископаемых пдауновид- ных было немало форм, способных к вторичному утолщению (рис. 230). Ныне живущие формы такой способностью не обладают, поэтому в течение всей жизни растения стебель остается травянистым и не одревесневает. Сам побег чаще всего бывает ползучим или стелющимся, у многих видов он вьющийся (лианы), лишь немногие виды имеют относительно прямостоячий побег.


Рис. 230. Лепидодендровые:
I - лепидодендрон (Lepidodendron):
A – молодое растение;
Б - взрослое растение;
В - схема поперечного среза ствола;
Г - схема поперечного среза листа;
Д - листовая подушка на продольном разрезе ствола;
Е - листовая подушка в плане;
Ж - стробил; 1 - кора; 2 - луб; 3 - сердцевина; 4 - первичная древесина; 5 - вторичная древесина; 6 - язычок; 7 - парихны; 8 - листовой рубец; II - спорофиллы лепидодендровых;
9 – спорофилл с тонкостенным вскрывшимся спорангием, внутри которого находилось несколько спор; 10- мегаспорофилл с толстостенным спорангием, содержащим мегаспору;
II - мегаспорофилл с толстостенным спорангием, прикрытым крыльями пластинки спорофилла, внутри спорангия находится женский гаметофит (по Н. С. Снигиревской)
Размеры современных плауновидных обычно небольшие, они варьируют у представителей различных видов от нескольких сантиметров до нескольких десятков сантиметров. Однако некоторые тропические виды имеют прямостоячий побег, достигающий двухметровой высоты, а длина побега лиан может достигать 20 м.
Подавляющее большинство плауновидных обитает во влажных местах. Такая экологическая привязанность объясняется потребностью в капельно-жидкой воде при оплодотворении. При этом плауновидные обычно довольствуются очень незначительной освещенностью, занимая самый нижний ярус в лесу. Большинство селягинвлловых произрастают во влажных тропических лесах. Полушниковые ведут водный или полуводный образ жизни, при этом в высокогорных озерах они нередко доминируют над другими водными растениями, поскольку способны довольствоваться настолько малыми концентрациями углекислого газа и растворенных в воде минеральных веществ, что другие растения в подобных условиях просто не выживают. Плауны в основном также предпочитают хорошо увлаженные места, но в то же время успешно живут в разнообразных климатических условиях - от влажных тропиков до умеренных широт и даже в тундре или высоко в горах. Весьма оригинальными являются эпифитные формы, использующие в качестве опоры стволы деревьев. Кроме того, среди плауновидных имеются ксерофитные формы, способные выдерживать значительный дефицит влаги и произрастать на сильно освещенных местах.
Значение плауновидных в природе относительно невелико. Очень немногие из них поедаются травоядными животными, а содержащие сильнодействующее вещество, способное эффективно останавливать перенос нервного импульса через синапс (наподобие яда кураре), плауны вообще остаются без внимания, и лишь некоторые беспозвоночные изредка поедают их без вреда для себя.
Также скромно плауновидные используются в хозяйственной деятельности человека. В частности, заготавливаемые в значительных количествах споры плаунов применяются в медицине в качестве присыпки, ими пересыпают пилюли. Высокое содержание в спорах жира служит причиной использования их в изготовлении фейерверков и в металлургии. В медицине также нашли применение некоторые высокоактивные соединения, содержащиеся в плаунах. Некоторые виды полушников украшают аквариумы, а многие тропические селягинеллы, обладающие красивой окраской, охотно выращивают любители оранжерейных растений. В настоящее время на Земле насчитывается около 400 видов плауновых, 700 видов селягинелловых и 64 вида полушниковых.
Среди папоротникообразных членистые в настоящее время являются самым малочисленным классом. В него входит только один порядок — хвощовые с единственным семейством (хвощовые), к которому относится опять-таки единственный род - хвощ, насчитывающий 32 вида. Остальные виды вымерли в начале мезозойской эры и сохранились лишь в виде ископаемых остатков в залежах каменного угля.
Представители этого класса имеют выраженное расчленение побега на узлы и междоузлия (рис. 231). От узла наземного побега отходит мутовка веточек, которые часто несправедливо принимают за листья. Сами листья хвощовых в значительной мере редуцируются. Они, как правило, лишены хлорофилла и, срастаясь между собой, образуют влагалища, которые охватывают основания междоузлия. На верхнем крае влагалища сращение листовых пластинок нарушается и отдельные листья заметны в виде зубчиков (но иногда листья срастаются и там). У разных видов хвощей листья, образованные ими влагалища и зубчики неодинаковы по своей длине, форме и окраске, что весьма важно при определении видовой принадлежности.

Рис. 231. Хвощ полевой (Equisetum arvense):
I - летний бесплодный побег; II - весенний спороносный побег (1 - клубеньки, 2 - колоски с мутовками спорофиллов); III - спорофилл (спорангиофор) со спорангиями: А - вид сбоку; Б - вид снизу;IV - споры с развернутыми элатерами; V - спора с завернутыми алатерами; VI - схема прохождения пучков в стебле хвоща (по Н. А. Комарницкому и соавт.)
Поскольку листья хвощей практически не фотосинтезируют, эту функцию берет на себя стебель (разумеется, его освещенная надземная часть). Хлоренхима в нем находится в периферической части первичной коры (рис. 232). Там она более или менее последовательно перемежается с тяжами живых вытянутых клеток, оболочки которых содержат много кремнезема и поэтому обладают высокими механическими свойствами. При этом механические элементы соответствуют ребрам стебля, а хлоренхима располагается вдоль межреберных ложбинок.

Рис. 232. Хвощ полевой:
I - схема поперечного разреза хвоща в области междоузлия: Il - детальное строение стебля из участка X - Y схемы; 1 - кора; 2 - валекулярная полость; 3 - механическая ткань в ребрышках; 4 - ассимиляционная ткань в ложбинках; 5 - эндодерма; 6 - центральная полость стебля; 7 - проводящий пучок; 8 - центральный цилиндр; 9 - устьице; 10- эпидермис; 11 - основная ткань; 12- флоэма пучка; 13- метаксилема; 14- протоксилема; 15- центральная полость (по Н. А. Комарницкому и совет.)
Поскольку тяжи проецируются вдоль ребер стебля, они получили название каринальных (лат. carina - киль, в данном случае гребень, ребро). Относительно ложбинок стебля во внутренней части первичной коры находятся валекулярные полости, которые у молодого стебля заполнены водой, а у зрелого воздухом (полагают, что он играет важную роль в газообмене).
Проводящая система хвощей оригинальна. Она образована закрытыми коллатеральными пучками (отсутствие в пучке камбия исключает вторичный рост), ориентированными вдоль ребер стебля и соответственно вдоль каринальных тяжей. Вода по пучку транспортируется несколькими путями. Во-первых, по каринальным каналам. Эти каналы возникают в результате растворения элементов протоксилемы. Иногда в каналах можно обнаружить отдельные кольчатые или спиральные трахеиды протоксилемы, имеющие очень большую длину. Кнаружи от каринальных каналов находится флоэма, а по бокам от нее располагаются еще две группы латеральной метаксилемы, которую образуют лестничные трахеиды. По ним также движется водный раствор минеральных веществ.
В типичном случае осевой цилиндр снаружи окружен эндодермой. У некоторых видов эндодерма выстилает стель еще и изнутри или же каждый пучок в отдельности. Чрезвычайно интересно ведут себя проводящие пучки при переходе из одного междоузлия в другое. При этом незадолго до вышележащего узла пучок разделяется на три ветви. На уровне узла срединная ветвь отклоняется наружу и продолжается в лист, становясь его жилкой (листовой след), а боковые ветви сливаются с такими же боковыми ветвями соседних пучков и образуют пучок, который продолжается в вышележащее междоузлие. Листовые прорывы в осевом цилиндре в результате отхождения листовых следов не образуются.
Внутри от осевого цилиндра у молодого стебля располагается сердцевина, но у зрелого органа она разрушается и замещается наполненной воздухом полостью.
Поверхность стебля хвоща неровная и покрыта продольно ориентированными гребнями, или ребрами, между которыми располагаются ложбинки. Количество гребней возрастает по мере увеличения диаметра стебля. Оболочки клеток эпидермы пропитаны кремнеземом, что придает им высокую механическую прочность (напомним, что кремнеземом также пропитаны клетки механической ткани, тяжи которой располагаются под ребрами, а под тяжами находятся проводящие пучки, также ориентированные вдоль ребер и повышающие механическую прочность стебля). В эпидерме междоузлий стебля имеется большое количество устьиц, располагающихся продольными тяжами вдоль ложбинок между ребрами. Ширина тяжей может варьировать от узких цепочек до более широких скоплений. Устьица состоят из двух замыкающих клеток, регулирующих размер устьичной щели и расположенных над ними двух побочных клеток, оболочки которых пропитаны кремнеземом и на внешней стороне имеют небольшие выступы, которые конгруэнтны впадинам в оболочке соседней побочной клетки. Выступы одной клетки входят во впадины другой, образуя «замок», способствующий надежному замыканию устьичной щели.
Верхушечный рост хвощей обеспечивается апикальной меристемой, расположенной на конце побега. Там выделяется клетка, имеющая трехгранную форму (см. рис. 115-А). Ее последовательные деления по граням откладывают молодые узлы и междоузлия, при этом несколько слоев клеток, расположенных над узлом, долгое время сохраняют свойства интеркалярных (вставочных) меристем. Они защищены влагалищем расположенного ниже узла и определенное время продолжают делиться, в результате чего происходит рост побега в длину. Однако постепенно клетки вставочных меристем перестают делиться и трансформируются в специализированные ткани.
Под защитой листовых влагалищ находятся и почки, которые не всегда четко приурочены к пазухе листа (поэтому многие специалисты до сих пор не считают почки хвощей пазушными). Многие из них остаются в состоянии покоя (спящие почки), а другие продолжают развиваться в боковой побег, который постепенно прорывает листовое влагалище. Поэтому отдельные растения хвощей значительно различаются по степени разветвления своих побегов.
Строение подземных побегов - корневищ - во многом сходно с наземными, однако имеется ряд различий (рис. 233). Сами побеги имеют значительно более ровную поверхность по сравнению с наземными (ребра на них почти не выражены). Поскольку они не освещены, отсутствует фотосинтезирующая хлоренхима. Кроме того, у подземных побегов нет устьиц. Зато в клетках основной паренхимы могут накапливаться значительные запасы крахмала, которые по мере необходимости используются для нужд растения. Большинство закладывающихся почек не развиваются в боковые побеги, зато на них в виде мутовок образуются придаточные корни. Способность формировать корни имеется и у почек наземных побегов, потому что и зачаточные побеги, и корни развиваются из общей массы клеток, возникших из одной материнской. При этом из клеток верхней части этой массы развивается боковой побег, а из нижней - корень. Соответственно у наземных побегов преимущественно развивается побег (хотя если его присыпать почвой, корни появятся), у подземных - корень. Любопытно, что часть корней у хвощей относительно толстая и длинная, демонстрирует обязательный для корней вообще положительный геотропизм. Другая часть, более многочисленная, имеет меньшую длину и толщину и на силу тяжести не реагирует.
Некоторые почки трансформируются в клубеньки, представляющие собой отдельные укороченные междоузлия. В клетках клубеньков в очень большом количестве откладывается крахмал.

Рис. 233. Подземные органы хвоща (Equisetum):
I - участки горизонтального и вертикального корневища с клубнями у хвоща полевого (Equisetum arvense); Il - то же у хвоща болотного (Equisetum palustre); III - участок корневища с клубнями у хвоща большого (Equisetum telmateia); IV - схема поперечного среза клубня хвоща полевого; V - схема поперечного среза положительно геотропичного корня хвоща полевого; 1 - покровная ткань; 2 - проводящий пучок; 3 - воздушная полость (по В. Р. Филину)
Все современные хвощовые являются равноспоровыми растениями (разноспоровость имела место лишь у некоторых вымерших форм). Спорангии развиваются на особых, присущих только хвощовым, структурах - спорангиофорах. Побеги, на которых они образуются, у большинства видов могут наряду со спорогенной функцией выполнять и фотосинтетическую. Такие побеги имеют очень высокую механическую прочность, они вечнозеленые и многолетние. У других видов (например, у хвоща лесного, хвоща полевого и др.) зеленые однолетние побеги бывают двух типов - чисто вегетативные и спорогенные. На последних вначале образуются стробилы, но после созревания спор и опадания стробилов эти побеги нельзя отличить от обычных вегетативных. И наконец, у третьих (например, у хвоща полевого) однолетние побеги изначально четко дифференцируются морфологически и функционально. При этом имеющие большую толщину спорогенные побеги лишены хлорофилла и имеют розовато-бурую окраску. Они не ветвятся, появляются рано весной вместе с вегетативными побегами или несколько раньше их и отмирают в тот же год обычно сразу же после спороношения.
Стробилы у хвощей образуются на верхушке главного побега или (реже) боковых ветвей. У большинства видов стробилы единичны, но у хвоща многощетинкового их численность может достигать ста. Основание стробила окружено видоизмененным листовым влагалищем - воротничком, а на его оси собраны мутовки многочисленных спорангиофоров (напомним, что так у хвощей называются спорофиллы). Спорангиофоры имеют ножку, которая заканчивается щитовидным диском, чаще всего имеющим форму шестигранника. На нижней части щитовидного диска развивается до полутора десятков спорангиев, расположенных вокруг ножки спорангиофора.
Спорангии образуются из группы эпидермальных клеток, среди которых выделяется одна самая крупная - осевая клетка. В результате последовательных делений образуется клеточная масса, из которой наружные клетки впоследствии образуют стенку спорангия (она в начале развития многослойная, а у зрелого спорангия состоит из одного слоя клеток), а внутренние становятся споро- генными. Из спорогенных клеток в результате митоза образуются материнские клетки спор. Затем каждая из них делится мейозом, давая начало четырем (тетраде) одинаковым спорам. Внутри споры находится крупное ядро, окруженное небольшим количеством цитоплазмы с многочисленными хлоропластами. Оболочка спор устроена довольно сложно. Кроме обычных двух оболочек - интины и экзины (эндоспорий и экзоспорий), имеется еще и третья, расположенная снаружи (эписпорий). Эта оболочка не образует сплошного слоя, а трансформируется в две ленты, по спирали закрученные вокруг споры, - пружинки, или элатеры (см. рис. 231). Как и положено эла- терам, они гигроскопичны и, закручиваясь вокруг споры в сырую погоду, раскручиваются в сухую, способствуя тем самым рассеиванию спор.
Спорофиллы с созревающими спорангиями на оси стробила располагаются близко друг к другу, но по мере завершения процесса спорогенеза несколько отдаляются друг от друга (это осуществляется из-за деятельности интеркалярных меристем). Истонченная стенка зрелого спорангия разрывается продольной щелью (чему немало способствует давление, которое изнутри оказывают на стенку спорангия элатеры спор), и через нее выходят споры, но не по одной, а комочками, что также обеспечивается элатерами. Комочки спор подхватываются ветром и разносятся на значительные расстояния.
Подавляющее большинство спор погибает, оказавшись в неблагоприятных условиях. Поэтому, несмотря на то что они образуются в огромных количествах, лишь очень немногие продолжают развитие, попав на хорошо увлажненную почву, защищенную от прямого света. Прорастание споры начинается с деления ее ядра (рис. 234). Из образовавшихся при этом двух клеток одна трансформируется в первичный ризоид, а из другой развивается фотосинтезирующая часть заростка с половыми органами. Первоначально молодой гаметофит напоминает нитчатую водоросль, но со временем приобретает довольно сложно устроенную пластинчатую форму с многочисленными уплощенными зелеными выростами, направленными вверх. На нижней поверхности заростка формируются бесцветные нитевидные ризоиды, снабжающие растение водой и минеральными веществами. В зависимости от условий развития размеры заростков варьируются от миллиметра до нескольких сантиметров.

Рис. 234. Прорастание споры, строение заростков (гаметофитов), гаметангиев и зародыша у хвоща (Equisetum):
А - Г - прорастание споры (и первые стадии развития заростка у хвоща ветвистого (Equisetum ramosissimum): 1 - заростковая клетка; 2 - срединный слой; 3 - ризоидальная клетка, 4 - эпатеры;
Д - молодой заросток хвоща большого (Equisetum telmateia): 5 - пластинка; 6 - меристема подушки; 7 - ризоид; 8 - подушка;
Е - мужской заросток хвоща ветвистого: 9 - антеридиальная ветвь;
Ж - женский заросток хвоща ветвистого; 3 - обоеполый заросток хвоща ветвистого: 10- антеридиальная ветвь;
И - архегоний хвоща ветвистого: 11 - шейка архегония; 12- яйцеклетка;
К - антеридии хвоща ветвистого: 13- клетки крышечки; 14- сперматогенная ткань;
Л - схема движения спиральной волны по жгутику сперматозоида хвоща (по В. Р. Филину)
Несмотря на то что хвощовые являются равноспоровыми растениями, их гаметофиты далеко не всегда бывают однодомными. По причинам, которые пока не известны, на заростке в одних случаях могут образовываться или только антеридии, или только архегоний, а в других гаметофит однодомен. Мужские гаметофиты по размерам уступают женским, но половые органы на них появляются немного раньше. У обоеполых гаметофитов последовательно сменяют друг друга три стадии: женская (когда первыми появляются архегоний), обоеполая (при последующем возникновении антеридиев), мужская (когда после отмирания архегониев остаются одни антеридии). Вообще, при определенных условиях однополые гаметофиты способны изменять свой пол (причем женские заростки трансформируются значительно чаще мужских).
В погруженных в ткань гаметофита антеридиях развивается несколько сотен крупных многожгутиковых сперматозоидов. Архегоний также погружены в гаметофит, но над поверхностью возвышаются их шейки. Оплодотворение у хвощей идет обычным для споровых растений способом в присутствии капельно-жидкой воды. Зигота дает начало зародышу, который состоит из корешка, стебелька и двух-трех листочков. От других папоротникообразных зародыш хвощей отличается тем, что не имеет подвеска и необходимые для развития вещества он получает из гаметофита через основание первого листового влагалища. Вначале зародыш окружен тканью гаметофита, но по мере развития корешок пробивает ее и оказывается в почве, где начинает активно выполнять свои функции. Но зародышевый корешок живет недолго. Из первичного побега сначала образуется корневище (от узлов которых отходят придаточные корни), а затем надземные побеги.
Как и все папоротникообразные, хвощовые очень древние растения. Самые ранние ископаемые остатки предковых форм обнаружены в отложениях начала девонского периода палеозойской эры, но максимального развития и разнообразия форм хвощовые достигли в каменноугольном периоде той же эры. Тогда были широко распространены очень похожие на современные хвощи древовидные каламиты (рис. 235), достигавшие двадцатиметровой высоты, которые, наряду с другими крупными папоротникообразными, образовывали целые леса. После этого (в триасовом периоде мезозойской эры) настала эпоха более высоко организованных семенных растений, и споровые (хвощовые в их числе) в большинстве своем вымерли.
Современные хвощи не способны к вторичному утолщению стебля, поэтому древовидные формы в настоящее время отсутствуют. Наиболее крупные хвощи растут во влажных тропических лесах и способны достигать десятиметровой длины, однако такой стебель не способен поддерживать себя в пространстве и в качестве опоры использует более мощные растения. Подавляющее большинство видов имеет относительно скромные размеры - от нескольких сантиметров до метра в высоту. При этом следует отметить, что четко определенные размеры отнюдь не являются постоянной характеристикой вида, поскольку могут широко варьировать в зависимости от условий произрастания растения.

Рис. 235. Каламит (Calamites carinatus):
A - общий вид;
Б - поперечный разрез стебля; 1 - вторичная древесина; 2 - каринальная полость; 3 - метаксилема (по Н. А. Комарницкому и соавт.)
Несмотря на небольшое видовое разнообразие, хвощовые являются очень распространенными растениями во всех географических областях, за исключением Австралии и Новой Зеландии (хотя искусственным путем они в настоящее время занесены и туда). Эти растения легко приспосабливаются к разнообразным условиям произрастания, заселяя болота, луга, пашни и леса на многих широтах. Часто хвощи численно доминируют над другими растениями, поскольку с успехом растут на переувлажненных почвах или в непосредственной близости грунтовых вод. Хорошо развитая система подземных побегов позволяет им выживать после пожаров.
Значение хвощей в природе и хозяйственной деятельности человека невелико. Травоядные животные в естественных условиях не поедают большинство из них, а попадание хвощей в корма для домашних Животных (если поле, на котором заготовлялись сенопродукты, было сильно засорено хвощами) резко снижает их ценность, а в больших количествах может привести к гибели животных, поскольку они содержат токсичные вещества. Значительные трудности возникают при очищении полей от хвощей, где они являются сорняками (хорошо развитые корневища при культивировании многократно разрубаются, что способствует не уничтожению хвощей, а напротив, их вегетативному размножению). Молодые спороносные побеги и клубеньки, содержащие большое количество крахмала, во многих странах население употребляет в пищу. Некоторые соединения, обладающие биологической активностью, употребляют при изготовлении лекарств. Жесткие стебли хвощей используют для полировки различных материалов, а также для чистки металлической посуды.
Вопросы для самоконтроля и повторения
1. Какое поколение преобладает в жизненном цикле папоротникообразных?
2. Почему папоротникообразные могут произрастать не только в сырых местах, но и на открытых участках?
3. Назовите вегетативные органы спорофита.
4. Какой тип корневой системы имеется у папоротникообразных?
5. Где у папоротникообразных развиваются споры?
6. При каких условиях происходит оплодотворение у папоротникообразных?
7. Какие особенности имеют вегетативные органы папоротниковидных? Что такое вайи?
8. Какие функции выполняют листья папоротниковидных? Как на них располагаются спорангии?
9. Назовите равно- и разноспоровые папоротниковидные.
10. Как происходит развитие гаметофита у папоротниковидных?
11. Чем трофофиллы отличаются от спорофиллов?
12. Как развивается женский гаметофит из мегаспоры у разноспоровых плауновидных? Как развивается мужской гаметофит из микроспоры?
13. Чем отличаются вегетативные побеги хвоща от спороносных?
14. Как происходит развитие гаметофита у хвоща?