Само название этих растений говорит об их «водном» обитании. Однако следует уточнить, что далеко не все растительные формы, встречаемые в водоемах, действительно являются водорослями. Значительная часть их относится к высшим растениям и имеет все присущие им вегетативные (лат. vegetativus - растительный) органы, причем водный образ жизни ведут представители всех групп высших растений, кроме голосеменных. Однако все они являются вторичноводными, поскольку исторически (с точки зрения эволюционной теории) происходят от наземных форм, а в воду попали, осваивая новые места обитания. В отличие от них, водоросли никогда не прерывали своего водного существования.
Жизнь в воде предполагает сравнительно стабильные условия для большинства клеток, образующих тело организма. Все они почти одинаково освещены, вода, окружающая растение, предоставляет всем его частям одинаковый набор растворенных в ней веществ. Кроме того, вода обеспечивает температурный режим, сходный для всех клеток. В результате клетки водорослей не имеют между собой особых различий, а равные для всех условия отнюдь не стимулируют дифференцировку (лат. differentia - разность, различие) клеток в специализированные. Поэтому водоросли в большинстве своем вообще не имеют выраженных тканей (исключение составляют высокоорганизованные бурые водоросли, но и у них ткани малочисленны и слабо дифференцированы). Отсутствие тканей, в свою очередь, объясняет нерасчлененность тела водоросли на вегетативные органы. Таким образом, тело водоросли представляет собой единое слоевище (таллом, греч. thallos - зеленая ветвь), которое может иметь самую разнообразную рассеченность, но при этом оно не дифференцировано на вегетативные органы. Таким образом, тело водоросли представлено нерасчлененным на вегетативные органы слоевищем, или талломом.
Некоторые формы посредством ризоидов (греч. rhiza - корень, eidos - вид) прикрепляются к субстрату (зачастую очень прочно), но эти «корнеподобные» структуры отнюдь не являются корнями, а служат лишь для удержания водоросли на субстрате и противодействуют течению воды или волнам. В случае если часть таллома все-таки отрывается (к примеру, во время шторма), ризоиды, как правило, повторно не образуются, а оторвавшаяся часть водоросли будет плавать на поверхности или в толще воды, не очень при этом страдая. Часто плавающие фрагменты растений образуют весьма значительные массивы, мигрирующие по воле течений.
Морфологическое разнообразие водорослей огромно: от чрезвычайно простых одноклеточных до сложнорасчлененных, нередко визуально очень похожих на высшие растения со структурами, внешне напоминающими стебли, листья и даже плоды. Неодинаковы и размеры - от гигантского макроцистиса, достигающего в длину 60 м (рис. 1), до микроскопической хлореллы (рис. 2). Выделяют несколько основных групп водорослей, различающихся степенью сложности морфологической организации.
1. Амебоидная структура (греч. amoibe - изменение) присуща некоторым представителям золотистых (рис. 3), желто-зеленых и пирофитовых водорослей. Такие одноклеточные организмы не имеют жесткой оболочки и поэтому не способны сохранять постоянную форму тела. Их клетки способны образовывать отростки - псевдоподии, часто значительной длины. Отдельные организмы могут находиться изолированно от других, но иногда отростки сливаются или же объединяются сразу несколько клеток.
2. Монадная структура (греч. monas, monados - единица, неделимое) характерна для одноклеточных водорослей, имеющих жесткую оболочку или уплотненный поверхностный слой цитоплазмы и постоянную форму тела. Для перемещения тела в пространстве такие водоросли используют различное количество жгутиков. Некоторые виды вторично утрачивают жгутики, но при этом оболочка становится менее жесткой и позволяет клетке изменять свою форму, способствуя передвижению в воде. Некоторые представители имеют внутриклеточную структуру, способную реагировать на свет, - глазок, или стигму. Монадную структуру имеют очень многие представители зеленых (рис. 4), желто-зеленых, золотистых и пирофитовых водорослей, а также эвгленовых, если рассматривать их как растения.

3. Коккоидная структура (греч. kokkos - зерно) широко распространена среди одноклеточных форм с жесткой оболочкой и постоянной формой тела (рис. 5). Морфологически она определяется отсутствием каких- либо органоидов, обеспечивающих активное движение клетки - псевдоподий или жгутиков. Обычно такие клетки пассивно переносятся током воды. При этом для увеличения парусности многие формы образуют на поверхности причудливые выросты. Часто такие водоросли образуют колонии, погруженные в слизь или же без нее.
4. Пальмеллоидная структура представляет собой постоянное или временное объединение нескольких отдельных коккоидных клеток в общую слизистую массу (рис. 6). Часто такие колонии имеют значительные размеры и прикрепляются к субстрату.

Рис. 4. Монадная структура у зеленых водорослей: 1 - одиночная клетка Chlamydomonas; 2 - колония Pyrobotrys, образованная срастанием клеток; 3 - колония Eudorina, образованная слизью (по М. М. Голлербаху)
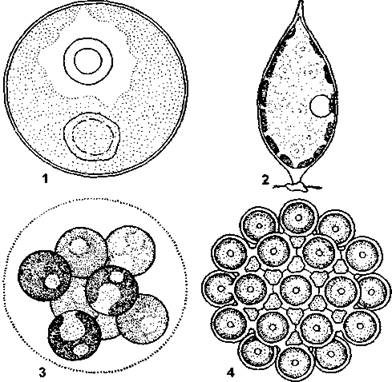
Рис. 5. Коккоидная структура у зеленых водорослей: 1 - одиночная клетка Chlorococcum; 2 - одиночная клетка Characium; 3 - колония Sphaerocystis, образованная слизью; 4 - колония Coelastrum, образованная срастанием клеток (по М. М. Голлербаху)
Рис. 6. Пальмеллоидная структура и пальмеллевидное состояние: 1 - пальмеллоидная структура у золотистой водоросли Hydrurus (часть таллома): 2 - пальмеллевидное состояние у зеленой водоросли Chlamydomonas (по М. М. Голлербаху)
5. Нитчатая структура является переходной формой к многоклеточной организации и среди водорослей чрезвычайно распространена. В этом случае тело растения представляет собой одиночную или разветвленную нить (рис. 7), которая ведет свободный или прикрепленный образ жизни. Деление клеток происходит лишь в одной плоскости, поэтому слои из клеток не образуются. Клетки в нити не имеют жгутиков и часто бывают связаны между собой плазмодесмами (см. раздел, посвященный строению растительной клетки). В наиболее простых случаях клетки в нити мало различаются между собой, но встречаются водоросли, где прослеживается полярность. При этом нижняя (базальная) клетка видоизменяется в ризоид, служащий для прикрепления к субстрату (рис. 8). Ближе к верхнему концу клетки по форме могут несколько отличаться от нижележащих. Деление может происходить либо во всех клетках нити (такой рост называют диффузным - лат. diffusio - распространение, растекание), либо в определенных зонах роста (своеобразных предшественниках меристем). Если такая зона находится в средней части таллома, рост водоросли называют интеркалярным (лат. inter - между), в верхней части - апикальным (лат. apex - вершина), а в нижней - базальным (грек, basis - основание). Если все нити ориентированы одинаково, структурная организация называется равнонитчатой, если часть нитей стелится по субстрату, а другая часть направлена вертикально - разнонитчатой (рис. 9).
Рис. 7. Нитчатые водоросли: 1 - улотрикс (Ulothrix zonata); 2 - стигеоклониум (Stigeoclonium tenue) (по «Жизнь растений», т. 3, с изменениями и дополнениями)
Рис. 8. Хризотриховые водоросли: 1 -4 - нитчатый таллом Nematochrysis scssilis (1 - зооспора, 2 - прорастание зооспоры, 3 - средний участок таллома, 4 - нижний участок таллома с ризоидальной клеткой); 5 - кустистый таллом Phaeothamnion confervicola (по А. М. Матвиенко)
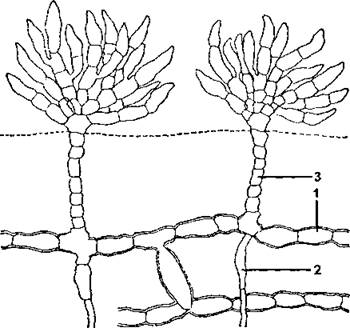
Рис. 9. Строение хетофоровых водорослей (Fritschiclla tuberose): 1 - стелющиеся нити; 2 - ризоиды; 3 - вертикальные нити (по К, Л. Виноградовой)
6. Пластинчатая структура происходит из нитей, в которых веретено деления располагается в различных плоскостях, в результате чего возникает более сложная организация клеточных ансамблей. При этом деление клеток в двух плоскостях приводит к образованию однослойной пластинки (рис. 10), если клетки делятся в трех взаимно перпендикулярных плоскостях - возникает многослойная пластинка (рис. 11). Именно пластинчатая структура по праву может считаться истинно многоклеточной.

Рис. 10. Пластинчатая структура у зеленой водоросли разиолы (Rasiola): однорядная нить, разрастающаяся в однослойную пластинку (no М. М. Голлербаху)
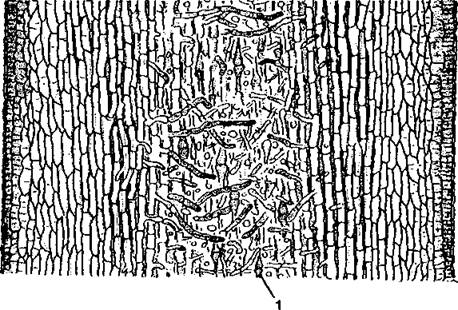
Рис. 11. Продольный разрез молодого слоевища ламинарии (Laminaria) при малом увеличении: 1 - трубчатые нити (по Ю. Е. Петрову)
7. Сифональная структура (греч. siphon - трубка), пожалуй, является самой оригинальной. В этом случае весь таллом образован всего лишь одной гигантской клеткой. Следует отметить, что в протопласте клетки содержится множество ядер, поэтому такую организацию можно объяснить тем, что после деления ядер не происходит образования поперечных перегородок и непосредственно сама клетка не делится. Внешний вид таких водорослей может быть весьма разнообразным - в виде ветвящихся нитей, Шаров и т. д. (рис. 12).

Рис. 12. Сифоновые водоросли: 1 - бриопсис: а - общий вид, б - верхняя часть ветви с гаметангиями; 2 - кодиум: а - общий вид, б - часть поперечного разреза; 3 - вапония; 4 - каулерпа (общий вид): а - Caulerpa macrodisca; б - Caulerpa proliféra; в - поперечный разрез горизонтальной части таллома (no Н. А. Комарницкому и соавт.)
Как уже говорилось, водоросли, за исключением некоторых просто устроенных наземных представителей, постоянно живут в воде и никогда ее не покидают. Здесь можно обнаружить бентосные (греч. benthos - глубина), или донные, формы, которые ризоидами прикрепляются ко дну, подводным камням, скалам или затонувшим предметам. Кроме этого, многие водоросли свободно плавают в толще воды, причем самые мелкие из них входят в состав фитопланктона (греч. phyton - растение, plankton - блуждающее) - продуцентной основы большинства водных биоценозов (рис. 13). Являясь фототрофами, водоросли нуждаются в солнечном свете, который поставляет необходимую для фотосинтеза энергию. Поскольку свет постепенно задерживается в толще воды, их распространение на глубину строго лимитировано. Это обстоятельство еще больше осложняется с увеличением мутности воды - в этом случае глубина заселения водорослями не превышает нескольких десятков сантиметров. В прозрачных водах морей и океанов водоросли можно встретить на глубинах, превышающих 150 м. Далее свет проникает в количествах, недостаточных для синтеза органических веществ, и фототрофные организмы здесь отыскать уже невозможно. В итоге, несмотря на огромную площадь поверхности планеты, покрытой Мировым океаном, бентосные водоросли можно обнаружить лишь на сравнительно узкой прибрежной полосе и на возвышениях дна.

Рис. 13. Внешние приспособления к планктонному образу жизни у водорослей из разных систематических групп:
1-4- шиповатые формы (1 - Mallomonas, одноклеточный жгутиконосец из золотистых водорослей с окремнепыми чешуйками на оболочке, снабженными отростками, 2 - колониальная зеленая водоросль ледиаструм (Pediastrum) с шипами на краевых клетках, 3 - одноклеточная зеленая водоросль голенкиния (Golenkinia) с шипами, усеивающими оболочку, 4 - одноклеточная диатомея коретрон (Corethron) с тремя венчиками отростков на панцире); 5 - 6 - парашютные формы (5 - звездчатая колония диатомеи астерионеллы (Asterionelia) со слизистыми тяжами между клетками, образующими парашют, 6 - одноклеточная диатомея планктониелла (Planktoniella) с плоской формой панциря) (по И. И. Николаеву)
То обстоятельство, что свет, проходя через толщу воды, постепенно рассеивается, вносит существенные коррективы в соотношения фотосинтетических пигментов. Значительно возрастает роль вспомогательных пигментов (более подробно об этом рассказано в разделе, посвященном объяснению фотосинтеза). При этом основной фотосинтетический пигмент - хлорофилл - может быть ими замаскирован. В результате вместо характерной для растений зеленой окраски водоросли могут приобретать и другие цвета, причем соответствующий набор пигментов зависит от глубины обитания организма. Этим воспользовались систематики, разделившие большинство водорослей на отделы в соответствии с набором их фотосинтетических пигментов. Систематическое положение водорослей неоднократно менялось, и в настоящее время наиболее приемлемо деление на следующие отделы. Напомним, что отделом у растений называется таксономическая, т.е. систематическая, категория, соответствующая у животных таксону тип, а порядок - таксону отряд.
1. Отдел Золотистые водоросли представляет собой довольно древнюю группу водорослей (обнаружены в ранних отложениях палеозойской эры). Большинство из них одноклеточные, встречаются колониальные и истинно многоклеточные формы (рис. 14). Окраска хроматофоров - от золотисто-желтой до зеленовато-бурой, зависит от соотношения пигментов, из которых обнаружены хлорофилл а и различные каротиноиды (из которых следует выделить золотистый фукоксантин и желтый лютеин). Интересно, что вместо крахмала их клетки запасают липиды в виде капель и гранулы особого полисахарида - лейкозина. Несмотря на повсеместное распространение, большинство золотистых водорослей живет лишь в чистых водоемах с пресной водой. Обильное размножение приводит к «цветению» воды. Известно около 500 видов.

Рис. 14. Основные типы структуры тела золотистых водорослей:
1 - амебоидная (Brehmiella chrysohydra); 2 - амебоидная (Chrysarachnion insidians); 3 - монадная (Synochromonas pallida); 4 - коккоидная (Chrysobotrys parvula); 5 - пальмеллоидная (Hydtvrus foetidus); 6 - разнонитчатая (Phaeothamnion borzianum); 7- пластинчатая (Phaeodermatium rivulare); 8 — колониальная (Dinobryon) (no «Жизнь растений», m. 3 и H. А. Комарницкому и совет.)
2. Отдел Желто-зеленые водоросли представлен всеми основными морфологическими формами (рис. 15). Хроматофор окрашен в различные оттенки желтого цвета благодаря наличию большого количества каротиноидов (эта группа пигментов количественно преобладает над другими). Кроме того, обнаружены хлорофиллы а и с. Трофические включения представлены липидными каплями, а также гранулами волютина и лейкозина. Эти водоросли распространены во всех водах и в почве, но большая часть живет в чистых пресных водоемах.

Рис. 15. Желто-зеленые водоросли:
1-3 — ботридиум (Bolrydnim grtimihnum) (1 - поросль на почве при малом увеличении, 2 - отдельная особь при большом увеличении, 3 - образование ризоцист); 4 - мисхококкус (Mischococcus confervicola); 5 - хлоротециум (Chlorothecium crassiapes), клетка и выход из нее зооспор; 6 - перонелла (Peroniella curvipes) (по «Жизнь растений», т. 3)
3. Отдел Зеленые водоросли характеризуется чрезвычайно широким морфологическим разнообразием своих представителей. У всех них хроматофор имеет выраженную зеленую окраску, она обеспечивается преобладанием над другими пигментами хлорофиллов а и в. Кроме них обнаружены различные каротиноиды. В качестве источника энергии запасается крахмал, иногда липиды. Большая часть зеленых водорослей живет в пресных водоемах, но есть и морские формы, а также почвенные. Это наиболее крупный по числу представителей отдел водорослей - сюда относят до 20 000 видов.
4. Отдел Красные водоросли, или багрянки, в подавляющем большинстве представлены нитчатыми или истинно многоклеточными формами. Морфологическое расчленение таллома может быть очень разнообразным и часто напоминает вегетативные органы высших растений (рис. 16). Среди бентосных водорослей представители именно этого отдела распространены наиболее широко. Размеры таллома колеблются от микроскопических до 2 м в длину. Окрашенные в красный цвет хроматофоры иногда называют радопластами. Их цвет определяется сочетанием хлорофиллов а, в и d (который больше не встречается ни у каких растений), каротиноидов и особых пигментов - фикобилинов. Высокое содержание красных пигментов позволяет красным водорослям заселять значительные глубины, не доступные большинству других водорослей (при высокой прозрачности воды они могут заселять донные субстраты на глубинах до 100 и даже 200 м). Запасным веществом служат липиды и особый полисахарид- багрянковый крахмал, химически занимающий промежуточное положение между крахмалом и гликогеном. Оболочка клеток поверх целлюлозного слоя покрыта довольно толстым слоем пектинов, из которого получают агар-агар. Для красных водорослей характерно наличие весьма сложного цикла развития. За исключением небольшого количества пресноводных форм, большинство багрянок живут в водоемах с соленой водой. В отделе насчитывается около 4000 видов.

Рис. 16. Красные водоросли:
1 - одонтапия (Odonthalia ochotensis); 2 - токидодендрон (Tokidodendron bullata): 3 - родомепа (Rhodomela lycopodioides); 4 - дазия (Dasia baillouviana) (no «Жизнь растений», т. 3)
5. Отдел Бурые водоросли представлен исключительно многоклеточными формами. Все они входят в состав бентоса, прикрепляясь к субстрату с помощью ризоидов или расширенного основания таллома - базального диска. Размеры их весьма разнообразны, они варьируют от нескольких миллиметров до 60 м в длину (см. рис. 1). Внешнее строение слоевища также очень различно и нередко расчленено до такой степени, что легко может быть спутано с настоящими вегетативными органами высших растений (рис. 17). При этом образуются ветвящиеся «кусты», на ветках которых имеются похожие на листья уплощения, а воздушные пузыри, удерживающие тяжелые ветви водоросли в плавучем состоянии, напоминают плоды.

Рис. 17. Бурые водоросли:
А - пессония (Lessonia);
Б - вершина ветви хормозиры Банкса (Hormosira banksii);
В - егрегия (Egregia);
Г, Д- химанталия удлиненная (Himanthalla elongata): Г- молодое растение, Д - растение с рецептакулами;
Е - вершина ветви цистозейры бородатой (Cystoseira barbata);
Ж- вершина ветви цистозейры косматой (Cystoseira crinita);
З - вершина ветви саргассума (Sargassum);
И - вершина ветви турбинарии (Turbinaria); 1 - базальный диск; 2 - рецептакулы (по Ю. Е. Петрову)
Внутреннее строение таллома бурых водорослей также выделяет их в ряду низших растений. Прежде всего это относится к дифференцировке клеток на более или менее выраженные типы, что позволяет говорить о появлении у них нескольких типов тканей (до четырех - проводящие, механические, ассимиляционные и запасающие). Клетки в них связаны между собой посредством плазмодесм. В клетках бурых водорослей хроматофор обычно имеет фрагментарное строение, причем его строение позволяет делить организмы на группы. Подбор пигментов определяет окраску слоевища в разные оттенки бурого цвета. Здесь имеются хлорофиллы а и с, а также различные каротиноиды. Питательные вещества запасаются в виде капель жира, растворимого многоатомного спирта маннита и растворимого полисахарида ламинарина (запасание энергетически ценных веществ в живых клетках в виде настоящих растворов довольно необычно). В состав оболочек клеток бурых водорослей входит особый тип целлюлозы, который больше нигде в растительном мире не встречается. Эти водоросли живут в соленых водах, а некоторые представители имеют серьезное промышленное значение (например, бурая водоросль ламинария, или морская капуста, является важным пищевым продуктом). В отделе насчитывается около 1500 видов.
6. Отдел Эвгленовые водоросли представлены исключительно одноклеточными (рис. 18) или колониальными формами. Обычно округлые или вытянутые клетки не имеют целлюлозной оболочки, поэтому постоянную форму тела им позволяет сохранять пелликула - уплотненный слой периферической цитоплазмы и над мембранные структуры. У некоторых форм пелликула отсутствует или истончена, что позволяет клетке обратимо изменять свою форму.

Рис. 18. Зеленые эвгленовые водоросли:
1 - Euglena acus; 2 - Euglena convolute; 3 - Euglena gracilis; 4 - Phacus monilatus; 5 - Phacus orbicularis; 6 - Phacus iongicauda; 7 - Phacus arnoldii; 8- та же клетка сверху; 9 - Euglena oxyuris; 10 - Eutreptia pyrenoidifera; 11 - Lepocinclis marssonii; 12 - Colacium arbuscula (по T. А. Сафоновой, с изменениями и дополнениями)
Характерно наличие одного или двух жгутиков (они помогают в плавании и анимальном питании) и светочувствительного глазка - стигмы. В хроматофоре присутствуют хлорофиллы а ив, обеспечивающие зеленую окраску, а также каротиноиды. Эвгленовые водоросли живут в основном в мелких пресных и слабосоленых водоемах. В отделе по разным источникам насчитывается от 400 до 1000 видов. Поскольку данная группа организмов объединяет в себе признаки как растений, так и животных, их традиционно изучают и в курсе ботаники, и в курсе зоологии, поэтому более подробно о них будет рассказано в разделе, посвященном животным.
7. Отдел Харовые водоросли (лат. chara - дикая капуста, полевой тмин) очень своеобразен. Внешне эти водоросли очень напоминают хвощи своим расчлененным на узлы и междоузлия слоевищем (рис. 19). Причем это сходство настолько велико, что специалисты часто называют соответствующие части таллома «стеблями» и «листьями», хотя, конечно, никакого отношения к настоящим вегетативным органам высших растений они не имеют. Харовые водоросли довольно крупные (до метра длиной и даже более) растения. Они имеют вид ветвящихся кустиков, на которых вытянутые междоузлия метамерно сменяются мутовками укороченных боковых побегов, которые и получили название «листьев». «Стебли» обладают способностью к неограниченному верхушечному росту. К субстрату (чаще всего это донный ил) харовые водоросли прикрепляются с помощью нитевидных ризоидов.

Клетки в слоевище неодинаковые. Междоузлие образовано одной гигантской (до нескольких сантиметров) клеткой, причем у некоторых харовых снаружи она еще покрыта слоем более мелких клеток - корой. Гигантские клетки междоузлий многоядерные, но это носит вторичный характер. Первоначально будущая клетка междоузлия содержит одно ядро, которое в процессе дифференцировки многократно делится. Сама клетка при этом удлиняется, однако поперечные перегородки в ней не образуются и все ядра находятся в одном протопласте.
Такие клетки уже не способны делиться, но такую возможность некоторое время сохраняют клетки, расположенные в междоузлиях. Плоскость деления у них проходит не поперек продольной оси «стебля», а параллельно ей, при этом на боковой поверхности «стебля» образуется бугорок (рис. 20). Поскольку все это радиально симметрично, в итоге образуется мутовка «листьев» (а у многих еще и «прилистников» - рис. 21) и боковых «побегов».
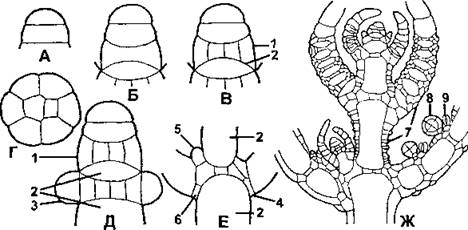
Рис. 20. Строение верхушки стебля хэрофитов в продольном сечении (схематизировано):
А- Е- схема начальных стадий верхушечного роста (А - двухклеточная стадия, Б - четырехклеточная стадия, В - начало формирования стеблевого узла и междоузлия, Г - поперечный разрез сформированного стеблевого узла, Д - начальная стадия образования листьев из периферических клеток стеблевого узла, Е - стеблевой узел с развитыми листьями и образующейся ветвью);
Ж - часть стебля хары на протяжении верхних четырех мутовок; 1 - стеблевой узел; 2 - междоузлие: 3 - начальные листовые бугорки; 4 - базальный листовой узел; 5 - начальный бугорок боковой ветви стебля; 6 - клетки, образующие прилистники; 7 - клетки стеблевой и листовой коры; 8 - антеридии; 9 - оогонии (по М. М. Голлербаху)

Рис. 21. Основания листовых мутовок разных видов хары с различно развитым венчиком «прилистников»:
А - однократный однорядный венчик Chara braunn;
Б - двукратный однорядный венчик Chara gymnopitys;
В-Д- двукратные двухрядные венчики (В - Chara fragilis, Г - Chara vulgaris, Д - Chara hispida) (по M. М. Гогтербаху)
Интересна структура зрелой клетки. Большую часть ее объема занимает крупная вакуоль, которая оттесняет к клеточной стенке цитоплазму с ядрами. Для харовых водорослей характерна очень высокая скорость движения цитоплазмы. Ее можно разделить на два слоя - внутренний, содержащий многочисленные ядра, и наружный, в котором находятся хлоропласты. За исключением неокрашенной полосы, пересекающей клетку вдоль ее продольной оси (рис. 22), хлоропласты очень равномерно располагаются в цитоплазме. Набор и соотношение фотосинтетических пигментов здесь такие же, как и у зеленых водорослей, поэтому и клетки харовых водорослей окрашены в равномерный зеленый цвет. В качестве запасного питательного вещества откладывается крахмал.

Рис. 22. Концевая часть листа нителлы (Nitella): левый членик - вид с поверхности; правый членик - вид в продольном разрезе (по М. М. Голлербаху)
Еще одной особенностью этих водорослей является их размножение. В отличие от других водорослей, у харовых имеются многоклеточные органы полового размножения - антеридии и оогонии (рис. 23, 24). Сформировавшиеся в оогониях яйцеклетки оплодотворяются мужскими гаметами - антерозоидами, которых в каждом антеридии образуется до 40 000. При этом образуется зигота. У харовых она интересна тем, что ее стенка пропитывается кремнеземом и суберином и затвердевает. Такое образование называется ооспорой, она имеет округлую или элипсоидную форму. Стенка ооспоры состоит из четырех слоев, два из которых окрашены в цвета от коричневато- желтого до черного. Внутри ооспоры запасаются питательные вещества - прежде всего крахмал и жировые капли. Какое-то время ооспора находится в состоянии покоя, после чего прорастает (рис. 25).

Рис. 23. Строение антеридия харофитов:
А - Г- последовательные стадии образования антеридия, вид с поверхности (А) и в продольном разрезе (Б - Г):
А - начальная восьмиклеточная стадия;
Б, В - последующие стадии обособления наружных щитовых клеток и внутренней части;
Г - поздняя стадия с дифференцированной внутренней частью;
Д - зрелый антеридий, вид с поверхности;
Е - два щитка с рукоятками, вид изнутри;
Ж - рукоятка отдельно с хорошо развитыми головками и антеридиальными нитями;
З - участок антеридиальной нити с антерозоидами в клетках и снаружи;
И - антерозоид при большом увеличении; 1 - исходная клетка основания; 2 - внутренняя базальная («бутылковидная») клетка; 3 - наружная базальная клетка; 4 - клетки листового узла; 5 - стенка; 6 - рукоятка, 7 - головки; 8 - антеридиапьные нити (no М. М. Голлербаху)
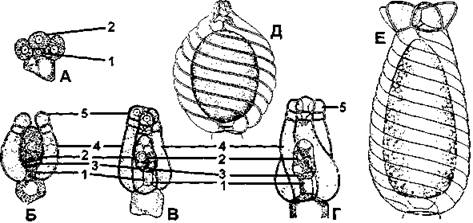
Рис. 24. Строение оогония харофитов:
А- Г- последовательные стадии образования оогония (А - начальная стадия, Б- Г- последующие стадии формирования яйцеклетки и обволакивающих спиральных клеток);
Д - зрелый оогоний нителлы (Nitella);
Е - зрелый оогоний хары (Chara); 1 - клетка листового узла; 2 - яйцеклетка; 3 - поворотные клетки; 4 - обволакивающие клетки; 5 - клетки коронки (по М. М. Голлербаху)

Рис. 25. Ооспоры и их прорастание:
А, Б- внешний вид зрелых ооспор нителпы (А) и хары (Б);
В-Д- последовательные стадии прорастания ооспор и формирования молодого растения хары (Chara canescens) (по М. М. Голлербаху)
Кроме того, на ризоидах и погруженных в грунт частях «стеблей» образуются органы вегетативного размножения - клубеньки (по расположению их часто называют корневыми и стеблевыми соответственно), которые могут быть одно- и многоклеточными (см. далее рис. 29).
Харовые водоросли обычно живут в чистых пресных водоемах со стоячей водой, где на дне образуют густые заросли. Реже их можно встретить в реках и еще реже в водоемах с солоноватой водой - в местах впадения рек в моря, чисто морских форм не существует. К отделу относят около 200 видов.
8. Отдел Диатомовые водоросли представляет собой одноклеточные, колониальные или нитчатые организмы (рис. 26) с жесткой клеточной оболочкой, содержащей большое - до 50 % массы клетки - количество кремнезема. Оболочка с кремниевым панцирем устроена очень сложно и может приобретать самые различные формы, в связи с чем их делят на две большие группы - радиально симметричные и билатерально симметричные. Подвижность клеток определяется наличием продольной щели в оболочке - шва. Полагают, что цитоплазма, двигаясь вдоль шва, создает трение о воду, что позволяет клетке двигаться в противоположном направлении. Клетки, лишенные шва, не способны произвольно перемещать свое тело в пространстве.
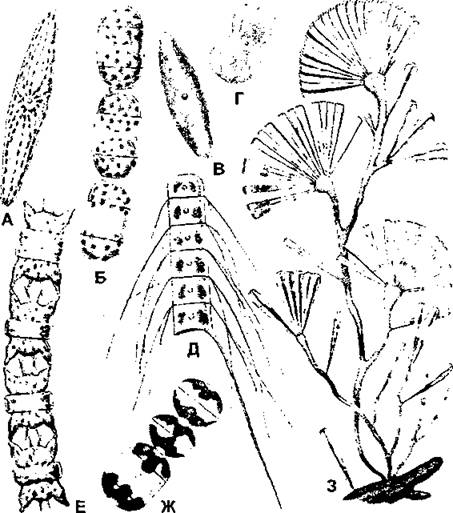
Рис. 26. Клетки диатомовых водорослей с хлоропластами различной формы:
А - плеуросигма (Pleurosigma sp.):
Б - мелосира (Melosira moniliformis);
В - навикула (Navicula sp.):
Г - амфипрора (Amphiprora sp.);
Д - хетоцерос (Chaetoceros subtilis var. abnormis);
E - биддупьфия (Biddulphia aurita);
Ж - гиалодискус (Hyalodiscus scoticus);
З - ликмофора (Licmophora sp.)
(по «Жизнь растений», m. 3)
Большая часть объема клетки заполнена одной или несколькими вакуолями, а цитоплазма с ядром занимает пристеночное положение. Хроматофоры живых диатомовых водорослей имеют желто-бурый цвет, что определяется пигментами - хлорофиллами а и с и каротиноидами. После гибели клетки происходит вымывание водой каротиноидов, и доселе замаскированный ими хлорофилл становится хорошо заметным, определяя посмертную зеленую окраску хроматофоров, а заодно и всей клетки. Запасными веществами является масло и полисахариды волютин и хризоза. Диатомовые водоросли заселяют любые типы водоемов, вплоть до термальных источников, погружаясь в глубину до 50 м. Входя в состав бентоса и планктона и обладая высокой энергетической ценностью, они являются важным компонентом водных экосистем. Отдел насчитывает более 10 000 видов.
9. Отдел Пирофитовые водоросли объединяет в основном одноклеточные формы, имеющие не радиальную, а билатеральную симметрию (реже асимметричны), в связи с чем для них характерно дорзовентральное строение тела (рис. 27). При этом обычно четко выделяются спинная, брюшная и боковые части, а также неодинаковы передний и задний концы клетки. Большинство передвигается при помощи жгутиков (рис. 28), длина и функции которых неодинаковы. Для многих характерно наличие сложно устроенной оболочки. Обязательным условием для пирофитовых водорослей является наличие бороздок, расположенных на брюшной стороне тела. Их может быть две или одна.
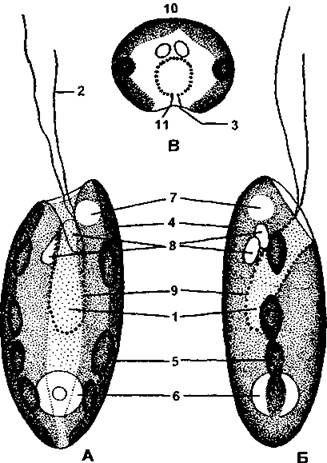
Рис. 27. Три положения клетки криптомонады:
А - клетка с брюшной стороны;
Б - клетка сбоку;
В - клетка сверху: 1 - клеточная глотка; 2 - жгутики; 3 - продольная бороздка; 4 - хлоропласты; 5 - пиреноид; 6 - ядро; 7 - сократительная вакуоль; 8 - тельца Monа; 9 - трихоцисты; 10- спинная сторона;
II - брюшная сторона
(по А. М. Матвиенко)
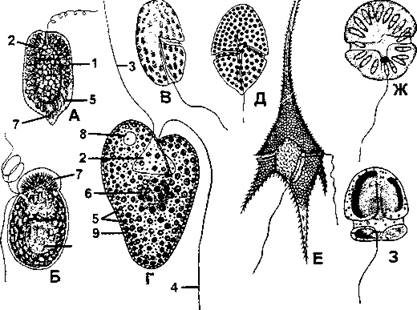
Рис. 28. Пирофитовые водоросли (хлоромонадофитовые):
А - гониостомум (Gonyostomum semen) - вид клетки со спинной стороны; Б - меротрихия (Merotrichia capitata); В - хемидимум (Hemidimum nasutum); Г - вакуопярия (Vacuolaria viridis) в поперечном разрезе; Д - гимниодиниум (Gymnodinium fuscum); Е - цератум (Ceratium hirundinella); Ж - гимниодиниум парадоксальный (Gymnodinium paradoxum); З - катодимум (Katodimum planmn): 1 - эктоплазма; 2 - клеточная глотка (цитофарингс); 3 - жгутик локомоторный; 4 - жгутик рулевой; 5 - хлоропласты; 6 - ядро; 7 - трихоцисты; 8 - сократительная вакуоль; 9 - слизистые тельца (по А. М. Матвиенко)
В клетке находятся несколько хроматофоров, содержащих пигменты: хлорофиллы а, с и каротиноиды. Соотношение этих пигментов обеспечивает удивительно разнообразную окраску хроматофоров - от оливкового до коричневого цветов. Запасными веществами являются липидные капли и зерна крахмала. Пирофитовые водоросли распространены повсеместно, они обитают во всех типах водоемов. Количество видов, входящих в отдел, достаточно велико - до 1100, но, как и в случае с эвгленовыми, на изучение этих организмов претендуют и зоологи, относя их к растительным жгутиконосцам в под царстве простейших.
Напомним, что, будучи прокариотами, синезеленые водоросли (цианобактерии) не могут быть отнесены к настоящим водорослям, поэтому здесь они не описаны.