Млекопитающие представляют собой наиболее высокоорганизованную группу высших позвоночных. У них лучше других развит головной мозг, особенно лобные доли. Для млекопитающих характерна сложная поведенческая деятельность, причем она основана не только на безусловных рефлексах (как у птиц), но и на условных, в связи с чем кора больших полушарий имеет очень большую площадь и у многих млекопитающих образует складки - извилины. Высокоразвитая условно-рефлекторная деятельность позволяет млекопитающим быстро приспосабливаться к изменяющимся условиям внешней среды, что делает этот класс самым процветающим среди других позвоночных.
Важным преимуществом является внутриутробное развитие плода, живорождение и вскармливание потомства молоком на начальной стадии постнатального развития. При этом млекопитающим присуща бережная забота о детенышах, они охраняют и обучают их, подготавливая к самостоятельной жизни. Для многих видов характерен коллективный образ жизни, что обеспечивает передачу информации от старых и наиболее опытных особей молодым.
Как и птицы, млекопитающие являются теплокровными животными, что позволяет им заселять обширные территории и занимать самые разные экологические ниши, при этом постоянно сохраняя высокий уровень активности, в том числе и в холодное время года. Всего насчитывается более 4500 видов млекопитающих.
Внешнее строение млекопитающих весьма разнообразно и зависит от образа жизни животного.
Анатомия. Наша задача во многом облегчается тем, что подробное строение наиболее совершенного вида млекопитающих - человека разумного - дано в первом томе настоящей книги. Поэтому здесь мы рассмотрим лишь общие черты строения и функционирования внутренних органов, по возможности избегая повторов.
Кожные покровы устроены сложно, что связано с их участием в терморегуляции. Ороговевающий эпидермис состоит из нескольких слоев, самый наружный - роговой, образованный мертвыми клетками, заполненными керато- гиалином, постепенно отпадает в виде чешуек или более крупных лоскутов.
Восстановление клеточной популяции идет за счет самого глубокого росткового (мальпигиева) слоя, в котором находятся постоянно делящиеся стволовые клетки. При делении стволовой клетки образуются две дочерние, которые не отличаются между собой по строению, однако имеют неодинаковую детерминацию. При этом одна из них детерминируется (направляется генетически) в функциональную клетку эпидермиса, и в последующем она пройдет все стадии дифференциации, после чего отомрет и отпадет в составе чешуйки. Другая клетка функционально не детерминируется и сохраняет свойства стволовой клетки, она никогда не будет дифференцироваться и остается «бессмертной», естественно, в пределах жизни самого организма (рис. 364).
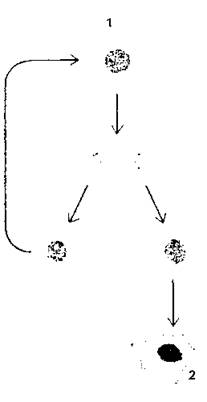
Рис. 364. Судьба потомков стволовой клетки.
Каждая дочерняя клетка, образующаяся при делении стволовой клетки, может остаться также стволовой, а может пойти по пути, ведущему к терминальной дифференцировке: 1 - стволовая клетка; 2 - терминально дифференцированная клетка (по Б. Албертсу и соавт.)
Из росткового (базального) слоя детерминированные клетки переходят в расположенный выше слой шиповатых клеток, которые имеют более крупные размеры и своеобразный внешний вид - они образуют многочисленные десмосомы с отходящими от них пучками кератиновых волокон (рис. 365), которые в световом микроскопе выглядят как шипы. Шиповатые клетки располагаются в несколько слоев. Кнаружи от них лежит более тонкий слой зернистых клеток, которые образуют границу в эпидермисе между внутренней зоной метаболически активных живых клеток и наружной зоной,
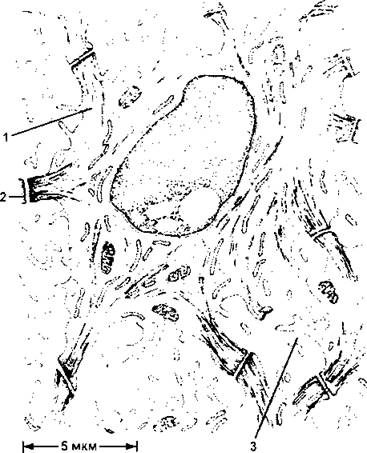
Рис. 365. Рисунок, сделанный по электронной микрофотографии среза шиповатой клетки эпидермиса. Видны пучки кератиновых нитей, которые пронизывают цитоплазму и направляются к десмосомам, соединяющим клетку с ее соседями. Обратите внимание, что между соседними клетками есть открытые каналы, позволяющие питательным веществам свободно диффундировать через метаболически активные слои эпидермиса. Ближе к его поверхности, на уровне зернистых клеток, имеется водонепроницаемый барьер, образованный, по-видимому, изолирующим веществом, который эти клетки выделяет из особых пузырьков:
1 - кератиновые филаменты; 2 - десмосома, соединяющая две клетки; 3 - открытый канал (по Р.В.Крстичу)
образованной мертвыми клетками, которые не содержат органоидов, а заполнены плотно упакованным кератином. Мертвые клетки образуют роговой слой, при этом клетки в нем настолько уплощены, что в световом микроскопе невозможно различить даже границ между клетками. Однако если эпидермис выдержать в растворе NaOH, то клетки набухнут и станут заметными колонки эпидермиса, демонстрирующие направление миграции детерминированных клеток в процессе дифференцировки (рис. 366). Такие колонки называются пролиферативными единицами эпидермиса.
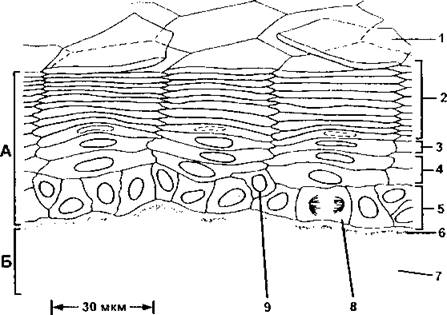
Рис. 366. Пролиферативные единицы, или колонки, в эпидермисе тонкой кожи. Эта структура выявляется при набухании ороговевших чешуек в растворе, содержащем NaOH. Такая организация в виде колонок свойственна лишь тонким участкам эпидермиса:
А - эпидермис; Б - дерма; 1 - чешуйка, готовая отделиться от поверхности; 2 - ороговевшие чешуйки; 3 - слой гранулярных клеток; 4 - слой шиповатых клеток; 5 - слой базальных клеток; 6 - базальная мембрана; 7 - соединительная ткань дермы; 3 - делящаяся базальная клетка; 9 - периферическая базальная клетка, переходящая в слой шиповатых клеток (по Б. Албертсу и соавт.)
Производными эпидермиса являются многие структуры: волосы, когти, ногти (отличаются от когтей тем, что они уплощены), копыта, рога (за исключением рогов оленей, которые имеют мезодермальное происхождение), чешуи, иглы. Кроме того, эпидермис образует различные кожные железы. Рога оленей целиком состоят из костной ткани, что отличает их от рогов других млекопитающих (рис. 367).
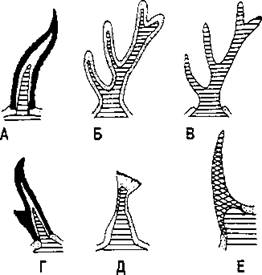
Рис. 367. Схемы рогов разных типов в продольном разрезе. Рога обращены передней стороной влево:
А - настоящий рог, такой, как у коровы или овцы; 6 - олений рог, покрытый кожей, когда рога завершают свой рост, эта кожа отмирает и отпадает; В - тот же рог без кожи; Г - рог вилорога; Д - рог жирафа; Е - рог носорога. На всех рисунках костный отросток черепа заштрихован, кожа покрыта точками, роговое вещество зачернено, а непарный рог носорога покрыт сеткой (по А. Ромеру и соавт.)
Волосы покрывают почти всю поверхность кожи, образуя характерный для млекопитающих волосяной покров, отсутствие волос у некоторых форм (например, ящеров, броненосцев, китов, дельфинов) носит вторичный характер. Различают несколько типов волос. Длинные, толстые и жесткие волосы - остевые - защищают кожу от повреждений, а также позволяют воде легко скатываться с тела животного, не смачивая его. Поэтому остевые волосы обычно располагаются к поверхности кожи под наклоном, чаще всего направленным назад и вниз, но у ленивцев, которые большую часть времени висят на ветвях дерева вниз спиной, волосы направлены от брюшной стороны к спинной и не препятствуют во время тропических ливней потокам воды стекать с тела. Пуховые волосы короче и мягче, у большинства млекопитающих они составляют основу волосяного покрова. Густой подшерсток задерживает вблизи кожи много воздуха, способствуя согреванию тела, поэтому пуховые волосы хорошо развиты у форм, обитающих в условиях длительных холодов. Специализированными волосами являются очень толстые и жесткие вибриссы, которые расположены на голове и выполняют функцию осязания (рис. 368).

Рис. 368. Схема расположения чувствительных полос (вибрисс) на морде кролика с очерченной пунктиром «осязательной зоной» (по С. П. Наумову)
Как и перьевой покров птиц, волосяной покров млекопитающих периодически меняется в ходе линек, которые обычно носят сезонный характер. Летняя и зимняя шерсть имеет неодинаковые свойства. В частности, зимний мех более густой, отдельные волоски длиннее, чем летом, кроме того, для многих млекопитающих характерна определенная сезонная окраска шерсти, обычно сходная с окружающим фоном, что позволяет животному быть малозаметным.
На дистальных фалангах пальцев большинства млекопитающих в зависимости от образа жизни развиваются когти, ногти или копыта. Уплощенные ногти характерны, в частности, для человекообразных обезьян, острые, загнутые когти необходимы лазающим животным и хищникам для удержания добычи. Копыта имеют то же происхождение, но они значительно мощнее и развиваются не на всех, а на двух (у парнокопытных) или одном (у непарнокопытных) пальце. Форма копыта у разных видов различна и зависит от условий жизни, например, если животное часто перемещается по мягкой почве или снегу, копыто широкое с большой площадью поверхности опоры для уменьшения давления на грунт, напротив, у горных копытных копыто узкое, что позволяет удерживаться на каменистых склонах.
Рога баранов, антилоп и быков также происходят из эпидермиса, они находятся на костной основе, сросшейся с лобными костями.
Для кожи млекопитающих характерно большое количество разнообразных желез эктодермального происхождения. Основными являются потовые и сальные железы. Потовые железы выделяют жидкость, состоящую из воды и растворенных в ней веществ (главным образом, неорганические соли и мочевина), компоненты секрета этих желез поступают в них из близлежащих кровеносных сосудов и тканевой жидкости.
Наряду с выделением продуктов обмена, другая главная задача потовых желез состоит в предохранении от перегревания тела, что основывается на уникальных свойствах воды. Напомним, что поляризованные молекулы воды (диполи) за счет взаимного притяжения посредством водородных связей образуют очень прочную молекулярную сеть. Для того чтобы молекулы смогли покинуть эту сеть, необходимо разорвать водородные связи (силы взаимного притяжения молекул воды, возникшие из-за разной электроотрицательности кислорода и водорода), что требует привлечения значительного количества энергии извне. Именно это свойство воды - высокая теплота испарения - лежит в основе охлаждения организма за счет пота. При этом жидкий пот, основу которого составляет вода, выделяясь на поверхность кожи, испаряется и отнимает у организма избыточное тепло, эффективно охлаждая его. При повышении температуры окружающей среды потовые железы функционируют более интенсивно, а при снижении количество выделяемого пота уменьшается.
Разумеется, процесс потообразования очень сложен и регулируется вегетативной нервной системой, которая оптимизирует количество и состав выделяемого пота в соответствии с нуждами организма. Особенно совершенна такая регуляция у видов, обитающих в суровых условиях регулярной засухи, например в пустыне. В частности, у верблюда при температуре +37°С на 100 см2 кожи испаряется 3 мг/мин, при +45°С - 15 мг/мин, а при +50°С - 25 мг/мин. Столь небольшое количество выделяемого пота позволяет верблюду экономить воду, поэтому животные, которые не привыкли к таким условиям, оказавшись в пустыне, быстро бы погибли от потери воды.
Количество потовых желез у разных видов неодинаково, например, их много у лошадей, но очень мало у собак и кошек и совершенно отсутствуют у китообразных (что в общем-то понятно, поскольку в водной среде испарение невозможно).
Сальные железы обычно открываются в волосяную сумку, их секрет покрывает тонким слоем волосы, придавая им водоотталкивающие свойства, кроме того, слой жира покрывает снаружи эпидермис.
И потовые и сальные железы могут видоизменяться. Часто пот имеет запах, причем характерный для определенного вида, кроме того, запах самцов отличается от запаха самок, поэтому секрет потовых желез имеет также важное значение в коммуникациях между особями, а хищникам позволяет легко отыскать добычу (во многом поэтому у них так хорошо развито обоняние). Иногда потовые железы, расположенные в определенных участках, видоизменяются в пахучие, которые выделяют сильно пахнущий секрет. Некоторые пахучие железы также являются видоизмененными сальными железами, иногда имеет место объединение в пахучих железах потовых и сальных желез. Лучше всего пахучие железы развиты у скунса, россомахи, ондатры, выхухоли и многих других животных.
Особенно важное значение имеют млечные, или молочные, железы, которые представляют собой видоизмененные потовые железы. Эти железы в полной мере развиты только у самок, причем функционируют они лишь определенное время после рождения детенышей (в период лактации). Выделяемый млечными железами секрет - молоко - служит основной (часто единственной) пищей на ранних стадиях постнатального развития детенышей млекопитающих, что и дало название всему классу. Выводные протоки млечных желез у большинства млекопитающих открываются на сосках, количество которых у разных видов неодинаково (например, у обезьян, овец, слонов и др. по два соска, а у мелких грызунов, насекомоядных и др. - до 24), но, как правило, это четное число. Только у однопроходных соски отсутствуют, при этом молоко выделяется на поверхность тела и детеныши его попросту слизывают.
Скелет млекопитающих имеет типичное для высших позвоночных строение (рис. 369). Позвоночный столб образован платицельными позвонками с плоскими сочленовными поверхностями (см. т. 1, рис. 142 и рис. 370), между которыми располагаются межпозвоночные диски. У млекопитающих четко различаются пять отделов позвоночного столба: шейный, грудной, поясничный, крестцовый и хвостовой. Шейный отдел у подавляющего большинства видов состоит из семи позвонков, лишь ламантин (шесть шейных позвонков) и различные ленивцы (от шести до десяти позвонков) являются исключениями. Первые два позвонка имеют особое значение, они образуют атлантоосевой комплекс, который обеспечивает подвижность головы. Затылочная кость черепа сочленяется с атлантом посредством двух затылочных мыщелков (напомним, что у земноводных также имеется два мыщелка, тогда как у рептилий и птиц - только один). Как и у птиц, поперечные отростки, срастясь с рудиментарными шейными ребрами, образуют характерные для шейных позвонков отверстия в поперечных отростках, через которые по всей длине шеи проходит позвоночная артерия, питающая головной мозг.
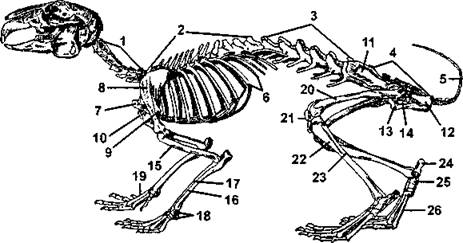
Рис. 369. Скелет кролика:
1 - шейные позвонки; 2 - грудные позвонки; 3 - поясничные позвонки; 4 - крестец; 5 - хвостовые позвонки; 6 - ребра; 7 - рукоятка грудины; 8 - лопатка; 9 - акромиальный отросток лопатки; 10- коракоидный отросток лопатки; 11 - подвздошный отдел безымянной кости; 12 — седалищный отдел той же кости; 13- лобковый отдел той же кости; 14 - запирательное отверстие; 15- плечо; 16- локтевая кость; 17 - лучевая кость; 18 - запястье; 19- пястье; 20 - бедро; 21 - коленная чашечка; 22 - большая берцовая кость; 23 - малая берцовая кость; 24 - пяточная кость; 25 - таранная кость; 26 - плюсна (по Брэму)
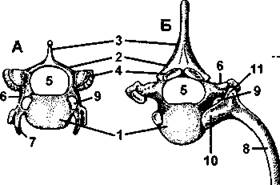
Рис. 370. Позвонки лисицы (вид спереди):
А - шейный позвонок; Б - грудной позвонок; 1 - тело позвонка; 2 - верхняя дуга; 3 - остистый отросток; 4 - сочленовные поверхности верхних дуг (для сочленения соседних позвонков); 5 - канал для спинного мозга; 6 - поперечный отросток; 7 - рудимент шейного ребра; 8 - ребро; 9 - отверстие для кровеносных сосудов; 10- головка ребра; 11 - бугорок ребра (по Н. Н. Карташеву и совет.)
Таким образом, длина шеи зависит не от количества позвонков, а от длины их тел. Наиболее длинная шея у копытных (вспомните жирафа!) и у хищных, для которых подвижная голова особенно важна. Между тем животные, ведущие подземный образ жизни (например, крот, цокор и др.), постоянно находятся в относительно узких норах, поэтому шея у них, как правило, короткая и малоподвижная.
Грудной отдел состоит из большего числа позвонков - от 9 до 24 у разных видов. Например, у клюворылого кита их 9, у лисицы - 13, а у ленивцев холоепусов - 24. Грудные позвонки отличает более крупное тело, но главное - к каждому из них присоединяется по паре ребер, состоящих из костного и хрящевого отделов. Каждое ребро своей костной головкой сочленяется с телом позвонка (см. рис. 370), а бугорком - с поперечным отростком. Костное тело ребра переходит в хрящевой отдел, который у передних ребер (истинные ребра) соединяется с грудиной, другие ребра до грудины не доходят (ложные ребра). Грудина представляет собой плоскую кость, состоящую из трех отделов: расширенного переднего - рукоятки, среднего - тела и заднего — мечевидного отростка, который обычно остается хрящевым.
Грудные позвонки, ребра и грудина в совокупности образуют хорошо выраженную грудную клетку, которая надежно защищает органы дыхания, сердце, крупные кровеносные сосуды и обеспечивает реберный тип дыхания. Жесткая грудная клетка делает грудной отдел позвоночного столба малоподвижным (особенно у крупных форм), этому также способствуют остистые отростки грудных позвонков, которые резко направлены назад и черепицеобразно накладываются друг на друга.
Поясничные позвонки наиболее мощные, они несут рудиментарные ребра, срастающиеся с поперечными отростками. Количество позвонков в этом отделе у разных видов млекопитающих варьирует от 2 до 9 (например, у лисицы их 7). Межпозвоночные диски здесь наиболее толстые, поэтому поясничный отдел у млекопитающих очень подвижен.
Крестцовые позвонки срастаются между собой, образуя единую кость - крестец, поэтому этот отдел неподвижен. Количество позвонков в крестце чаще всего 4, однако их может быть 3 (например, у хищных), 2 (у утконоса) или более 4 (до 10), но истинно крестцовыми являются только первые два позвонка, тогда как все остальные представляют собой хвостовые позвонки, сросшиеся с крестцом.
Численность хвостовых позвонков у разных млекопитающих широко варьирует. У длиннохвостого ящера 49 хвостовых позвонков, у кролика - их 15, у лисицы - 19, а у человекообразных обезьян все хвостовые позвонки срастаются в одну кость - копчик (например, у гиббона он образован тремя сросшимися хвостовыми позвонками).
Значение хвостового отдела для млекопитающих может быть весьма велико. Например, для паукообразных и некоторых других обезьян хвост выполняет функцию пятой конечности, в частности, животное может с помощью него висеть на ветке дерева, манипулируя при этом совершенно свободными конечностями, так же поступают древесный дикобраз и многие другие древесные млекопитающие. Для других хвост представляет собой балансир во время бега или прыжка, кенгуру во время прыжка использует свой мощный хвост в качестве опоры. В хвосте может откладываться жир, например у сусликов. Хвост белки покрыт тонкой шкурой, поэтому, если хищник хватает за него, кожа разрывается и шкурка, как чулок, соскальзывает с хвостового стержня, давая возможность зверьку спастись. Многие копытные отмахиваются хвостом от мух и других насекомых, а бегемот по не совсем понятным причинам быстрыми пропеллерообразными движениями короткого хвоста разбрызгивают свои экскременты в момент дефекации на большое расстояние. Для многих видов хвост позволяет выразить эмоции, например, собака, виляя хвостом, демонстрирует радость, а кошки, наоборот, размахивают хвостом в момент ярости.
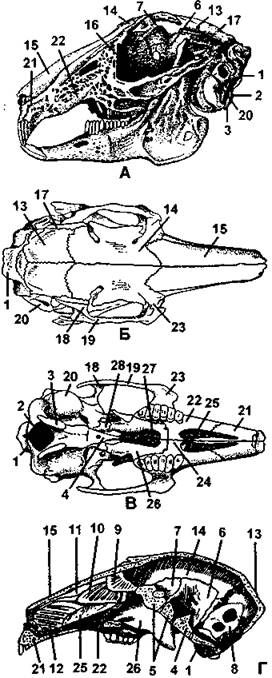
Рис. 371. Череп кролика:
I - сбоку; II - сверху; III - снизу; IV - сагиттальный разрез; I - затылочная кость; 2 - затылочное отверстие, 3 - затылочный мыщелок; 4 - основная клиновидная кость; 5 – переднеклиновидная кость; б - крылоклиновидная кость; 7 - глазокпиновидная кость; 8 - каменистая кость; 9 - решетчатая кость; 10 - обонятельные раковины; II - носовая раковина; 12 - челюстные раковины; 13 - теменная кость; 14- лобная кость; 15 - носовая кость; 16 — слезная кость; 17 - чешуйчатая кость; 18 - скуловой отросток чешуйчатой кости; 19 — скуловая кость; 20 - барабанная кость; 21 - межчелюстная кость; 22 - верхнечелюстная кость; 23 - скуловой отросток верхнечелюстной кости; 24 - нёбный отросток верхнечелюстной кости; 25 - сошник; 26 - нёбная кость; 27 - хоаны; 28 - крыловидная кость (по Паркеру)
Череп млекопитающих имеет ряд особенностей (рис. 371). Прежде всего он полностью окостеневает и у взрослых особей не содержит хрящевых элементов. Для млекопитающих характерно срастание отдельных костей черепа в более крупные костные образования, мозговой отдел срастается с висцеральным. В итоге череп млекопитающих, по существу, состоит из сплошного костного монолита, отдельные элементы которого соединены между собой только неподвижно (синхондрозы, которые постепенно заменяются синостозами, и швы). Исключением является нижняя челюсть, которая подвижно соединяется с височной костью мозгового черепа посредством височно-нижнечелюстного сустава. Подъязычная кость вообще непосредственно не сочленена с черепом и связывается с ним посредством надподъязычных мышц. Отдельную группу костей составляют слуховые косточки, расположенные в полости среднего уха. Еще раз напоминаем, что все остальные кости мозгового и висцерального черепа неподвижно срастаются между собой посредством непрерывных соединений.
Мозговой череп представлен парными и непарными костями. Затылочные кости большинства млекопитающих срастаются в единую кость, на ее наружной поверхности по бокам имеются два затылочных мыщелка для сочленения с первым шейным позвонком. Кроме того, в затылочной кости имеется большое затылочное отверстие.
Крышу черепа образуют парные лобные и теменные кости, которые являются накладными. Между затылочной и обеими теменными костями находится непарная межтеменная кость. Боковые стенки черепа формируются парными височными костями, которые возникли путем срастания чешуйчатой, каменистой и барабанной костей, превратившихся в ее отделы. Каменистая кость образуется сросшимися ушными костями, барабанная кость имеется только у млекопитающих, возможно, она гомологична угловой кости рептилий, в этой кости располагается среднее ухо. От чешуйчатого отдела вперед и кнаружи отходит скуловой отросток, который впереди соединяется со скуловой костью, та, в свою очередь, впереди соединяется со скуловым отростком верхней челюсти. Таким образом формируется характерная для млекопитающих скуловая дуга, которая является височной дугой смешанного типа, поэтому такой череп относится к синапсидному типу. Скуловая дуга ограничивает снаружи глазницу.
Основание черепа в задней части сформировано основной и боковыми частями затылочной кости, кпереди от нее располагается основная клиновидная кость, а впереди нее - узкая переднеклиновидная кость. Основная клиновидная кость по бокам срастается с крылоклиновидными костями, а переднеклиновидная также по бокам срастается с парной крыловидной, или глазоклиновидной, костью. У человека все перечисленные кости (кроме затылочной) срастаются в единую и очень сложную клиновидную кость. Впереди переднеклиновидной кости вертикально располагается непарная решетчатая, или обонятельная, кость, от боков которой отходят обонятельные раковины.
Висцеральный отдел черепа представлен верхней и нижней челюстями, а также подъязычным аппаратом. Верхние челюсти сформированы парными костями: межчелюстными, расположенными спереди, и значительно более крупными верхнечелюстными. Их нёбные отростки совместно с нёбными костями формируют твердое костное нёбо, которое разделяет носовую и ротовую полости млекопитающих. При этом хоаны открываются позади нёбных костей, что исключает попадание пищевых масс в носовую полость. Нижняя челюсть устроена еще проще - ее образуют только парные зубные кости, которые спереди неподвижно срастаются друг с другом. Напомним, что нижняя челюсть представляет собой единственную подвижную кость черепа млекопитающих, которая соединяется височно-нижнечелюстным суставом с чешуйчатой частью височной кости.
Характерные для нижней челюсти других позвоночных квадратная и сочленовная кости у млекопитающих находятся в полости среднего уха, где выполняют функции слуховых косточек - сочленовная кость видоизменилась в молоточек, а сочленовная - в наковальню. Таким образом, в среднем ухе млекопитающих находится не одна слуховая косточка (стремечко), как у всех остальных четвероногих, а три (молоточек, наковальня и стремечко), подвижно соединенные между собой суставами.
Подъязычный аппарат представлен подъязычной костью, состоящей из тела и двух пар рожек, из которых передняя пара гомологична нижней части подъязычной дуги, а задняя пара - первой жаберной дуге.
Подъязычная кость располагается между над- и подподъязычными мышцами и обеспечивает согласованные движения глотки и гортани.
Добавочный скелет имеет следующее строение. Пояс передней конечности (плечевой пояс) образован лопаткой и ключицей. Лопатка представляет собой плоскую кость с большой поверхностью для прикрепления мышц плечевого пояса. По дорзальной поверхности лопатки проходит ость, которая латерально переходит в вырост - акромион. Самостоятельный коракоид имеется только у однопроходных (утконоса, ехидны), но у всех остальных он срастается с лопаткой, образуя клювовидный отросток. Ключица имеется не у всех млекопитающих, а только у тех, у кого плечевой сустав является многоосным, т.е. способен совершать движения вокруг трех осей, например приматов, грызунов, зайцеобразных, рукокрылых, насекомоядных, сумчатых. Животные, у которых движения в плечевом суставе осуществляются преимущественно вокруг одной оси, ключицы не имеют, например у большинства хищных, хоботных, копытных, китообразных.
Свободная конечность имеет типичное строение пятипалой конечности: плечо образовано плечевой костью, предплечье - локтевой и лучевой, кисть состоит из запястья, пясти, фаланг пальцев. У летучих мышей все пальцы, кроме I, сильно удлинены, между ними натянута кожистая перепонка крыла.
Пояс нижних конечностей (тазовый пояс) у большинства млекопитающих состоит из парных тазовых, или безымянных, костей, соединенных на брюшной стороне тела посредством симфиза. Каждая из тазовых костей образуется в результате сращения трех костей: подвздошной, седалищной и лобковой. Поскольку лобковые кости, соединяясь между собой, образуют симфиз, таз у млекопитающих относится к закрытому типу.
Скелет свободной задней конечности состоит из бедра (бедренная кость), голени (большая и малая берцовые кости) и стопы (кости предплюсны, плюсны и фаланг пальцев). У млекопитающих локтевой сустав, соединяющий кости плеча и предплечья, направлен назад, а коленный сустав, соединяющий кости бедра и голени, - вперед (этот сустав укрепляет самая крупная сессамовидная кость - надколенник).
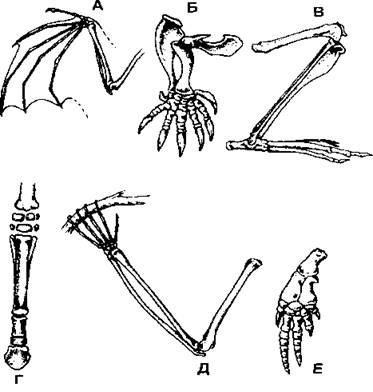
Рис. 372. Разнообразие строения конечностей млекопитающих: А - летучая мышь; Б - крот; В - кенгуру; Г-лошадь; Д - орангутан; Е - дельфин (по С. И. Левушкину и соавт.)
Несмотря на сходную общую схему строения свободных конечностей, разные виды имеют немало особенностей (рис. 372). Как уже упоминалось выше, пальцы кисти летучей мыши сильно удлинены, что связано с полетом. У водных млекопитающих дистальные отделы конечности также удлинены (кости пясти, плюсны и фаланги пальцев) при укороченных проксимальных отделах (плечо и предплечье, бедро и голень). Однако у наземных млекопитающих все наоборот - проксимальные отделы длиннее. При этом строение конечности зависит от образа жизни, например, у роющих видов конечность становится мощной и короткой. Млекопитающие, которые бегают относительно медленно, при ходьбе опираются на всю стопу (рис. 373), например медведи, обезьяны и др.
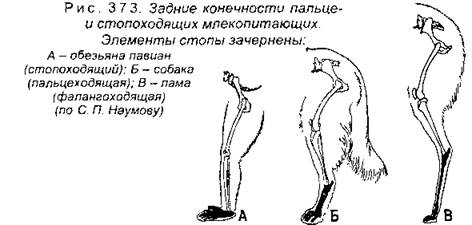
Более быстроногие звери опираются на пальцы, например собака, волк, лисица, шакал, кошачьи. Наконец самые быстрые бегуны опираются на фаланги пальцев, при этом количество пальцев у них уменьшается до двух - III и IV пальцы (у парнокопытных) или до одного - III палец (у непарнокопытных). Конечности млекопитающих располагаются под туловищем, а не по бокам его, как у пресмыкающихся, что позволяет поддерживать тело над поверхностью земли и делает перемещение более эффективным.
Мышечная система млекопитающих очень сложна, поскольку включает в себя большое количество четко дифференцированных мышц. Как и у птиц, наиболее мощные мышцы дистального отдела свободной конечности часто вынесены на более крупные кости предплечья или голени, тогда как к фалангам пальцев направляются лишь сухожилия (так, например, происходит у приматов). Мышцы пояса передней конечности прикрепляются к костям плечевого пояса и грудной клетки, мышцы пояса задней конечности прикрепляются, главным образом, к костям таза, а не хвостовых позвонков, как у пресмыкающихся. Важной особенностью скелетной мускулатуры млекопитающих является наличие у них куполообразной диафрагмальной мышцы, которая разделяет грудную и брюшную полости. Интересно, что эта мышца происходит от мышц шеи, но в период эмбрионального развития перемещается далеко назад, увлекая за собой кровеносные сосуды и нервы, обеспечивающие ее жизнедеятельность.
У млекопитающих хорошо развита подкожная мускулатура, иногда она образует под кожей сплошной слой (остатком такой мускулатуры являются прекрасно развитые мимические мышцы головы и малофункциональная подкожная мышца шеи человека). Подкожная мускулатура смещает участки кожи, поднимает шерсть (ощетинивание), иглы (у ежей и дикобразов), осуществляет движение вибрисс. Основа губ и щек образована подкожными мышцами.
Пищеварительная система млекопитающих отличается особой сложностью и многообразием частных особенностей, которыми обладает тот или иной вид. Ротовая полость ограничена спереди подвижными губами (они отсутствуют у однопроходных и китообразных), с боков - щеками, сверху - нёбом, снизу - диафрагмой рта, задняя стенка отсутствует и через широкое отверстие зева ротовая полость сообщается с глоткой. Для млекопитающих характерно разделение ротовой полости на преддверие (пространство между губами и зубами) и собственно ротовую полость (все остальное пространство), в которой находится язык. У грызунов, приматов и некоторых других млекопитающих преддверие ротовой полости настолько обширно, что образует защечные мешки. Однако не у всех млекопитающих губы располагаются впереди зубов, у некоторых видов (например, слепыша, слепушонка) все обстоит как раз наоборот - губы находятся позади зубов и предохраняют ротовую полость от попадания в нее комьев земли, когда животное прогрызает зубами свои норы в плотной почве (рис. 374).
В альвеолах верхних и нижней челюстей располагаются зубы (текодонтная зубная система), причем для млекопитающих характерна гетеро- донтностъ, т.е. имеет место морфологическое разделение зубов на функциональные группы. Различают резцы, имеющие долотообразный режущий верхний край коронки, клыки, с заостренной конической коронкой, предкоренные (малые коренные, ложные коренные, моляры), с жевательной поверхностью и бугорками на ней, коренные (моляры), у которых жевательная поверхность обширнее и больше бугорков.

Рис. 374. Голова гигантского слепыша (по С. П. Наумову)
Для разных видов млекопитающих характерен определенный набор зубов, графическая запись которого называется зубной формулой. В ней цифрами указывают количество зубов каждого типа на одной верхней челюсти и половине нижней челюсти, например, зубная формула волка:
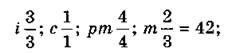
зубная формула кролика:
![]()
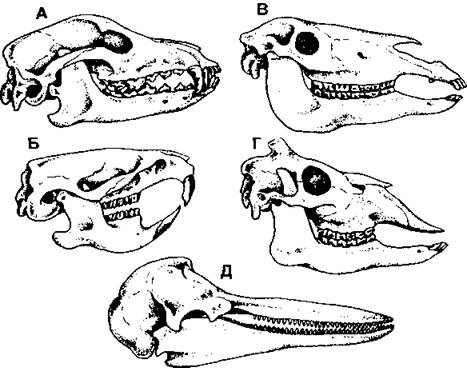
Рис. 375. Различные типы зубных систем млекопитающих: А - хищник; Б - грызун; В – непарнокопытное; Г – парнокопытное; Д - дельфин (по С. И. Левушкину и соавт.)
Набор зубов всегда зависит от образа жизни животного (рис. 375), при этом виды, сходные по типу питания, обычно имеют похожие зубные формулы и одинаковую форму зубов каждого типа (рис. 376). У млекопитающих смена зубов чаще всего происходит один раз в жизни - ранние (молочные) зубы сменяются постоянными (напомним, что у низших позвоночных, например акул, зубы меняются неограниченно часто). У грызунов резцы растут всю жизнь, поэтому животное вынуждено даже периодически грызть твердые предметы, чтобы сточить зубы до оптимальной длины.
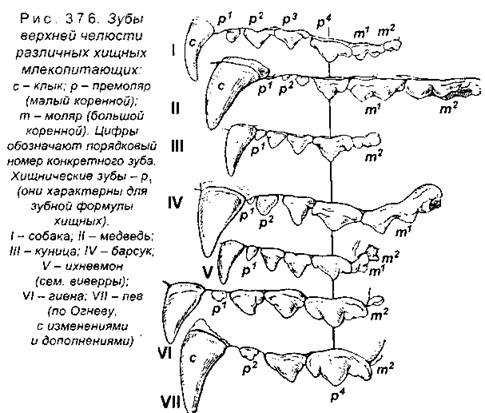
Малодифференцированные зубы имеют насекомоядные (землеройка, крот, выхухоль и др.) и неполнозубые (броненосцы, муравьеды, ленивцы), причем у последних зубы лишены эмали и корней или могут даже вообще отсутствовать. Зубы отсутствуют также у ящеров. У китообразных отмечается вторичное изменение зубов, при этом у зубатых китов (кашалотов), а также различных дельфинов они многочисленные, но одинаковые, а у усатых китов (синего кита, финвала, сейвала и др.) отсутствуют.
Дифференцированные зубы обеспечивают млекопитающим значительные преимущества перед другими классами позвоночных, поскольку позволяют им не только отрывать куски пищи, но и пережевывать их до гомогенной массы, что в значительной степени облегчает пищеварение и ускоряет процесс усвоения питательных веществ.
У млекопитающих хорошо развит язык, посредством которого животные схватывают пищу (например, бычьи, жирафы, муравьеды и др.), пьют (лакают), перемешивают пищевые массы в ротовой полости, передвигают пищевой комок в глотку и т.д. Длина языка очень широко варьирует, например, у жирафа он более полуметра, а у гуанако настолько мал, что животное не может даже как следует вылизать своего детеныша.
Слюнные железы у разных видов млекопитающих развиты неодинаково. Наиболее значительны они у жвачных (например, корова в течение суток выделяет до 56 л слюны), тогда как у китообразных вообще не развиты. Слюна млекопитающих содержит ферменты, расщепляющие углеводы (амилазу и мальтазу), поэтому в ротовой полости происходит не только механическая, но и частично химическая переработка пищи. У летучих мышей вампиров в слюне растворены антикоагулянты, препятствующие свертыванию в ранке крови, которой они питаются, слюна некоторых землероек токсична.
Глотка дифференцирована на три отдела: носоглотка (проводит только воздух), ротоглотка (проводит воздух и пищу) и гортаноглотка (проводит только пищу). Пищевод у млекопитающих хорошо развит, его длина соотносится с длиной шеи. Желудок всегда четко дифференцирован от соседних отделов пищеварительного тракта. У большинства видов желудок делится на несколько отделов, что связано с характером питания (рис. 377, 378).
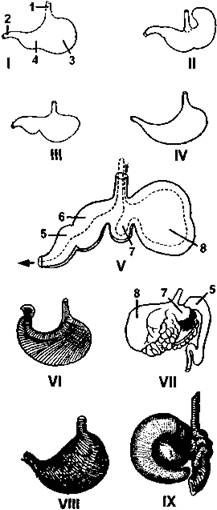
Рис. 377. Различные формы желудков млекопитающих:
I - собаки: II - крысы; III - мыши; IV - куницы: V - жвачного (пунктирная стрелка указывает направление движения пищи); VI - человека; VII - верблюда; VIII - ехидны; IX - трехпалого ленивца; 1 - пищевод; 2 - кишка; 3 - кардиальный отдел желудка; 4 - его пилорический отдел; 5 - сычуг; 6 - книжка; 7 - сетка; 8 - рубец
Наиболее сложно устроен желудок жвачных, состоящий из четырех отделов: рубца, сетки, книжки и сычуга (рис. 379). Пережеванная и пропитанная слюной пища из пищевода сначала попадает в рубец, имеющий на внутренней поверхности стенок твердые выпуклости, где происходит ее брожение под действием живущих там бактерий. Мышечные сокращения стенок желудка перемещают пищевую массу из рубца в сетку, стенки которой имеют вид ячеек. Из сетки пища отрыгивается обратно в ротовую полость (этому способствует поперечнополосатая мускулатура стенок пищевода), где повторно тщательно пережевывается, орошается слюной, после чего вновь проглатывается. Проглоченная полужидкая пищевая масса из пищевода попадает в книжку (ее стенки имеют продольные складки), а из нее в сычуг. Такое строение желудка позволяет жвачным перерабатывать очень грубые корма, кроме того, бактерии рубца перевариваются вместе с растительной пищей, обеспечивая животное полноценным белком, в частности, свиньи, которые не являются жвачными, вынуждены искать дополнительные источники белка (для этого они поедают трупы мелких животных, разоряют гнезда птиц и т.п.).
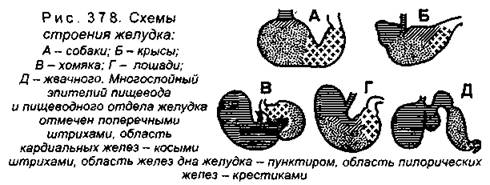
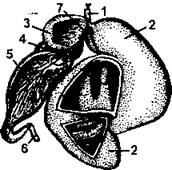
Рис. 379. Вскрытый желудок овцы: 1 - пищевод; 2 - рубец; 3 - сетка; 4 - книжка; 5 - сычуг; 6 - двенадцатиперстная кишка; 7 - желобок
Кишечник подразделяется на тонкую и толстую кишку, в тонкой кишке происходит расщепление полисахаридов (кроме целлюлозы), белков и жиров. В начальный отдел тонкой кишки - двенадцатиперстную кишку - открываются протоки печени (общий желчный проток) и поджелудочной железы (часто она не обособлена анатомической и диффузно располагается в толще брыжейки тонкой кишки), которые обычно предварительно объединяются в общий проток.
В толстой кишке происходит всасывание воды из переработанного химуса, формирование каловых масс и их накопление. У млекопитающих хорошо выражена слепая кишка, причем ее длина зависит от характера пищи, то же самое относится и ко всей толстой кишке в целом (рис. 380). Толстая кишка наиболее развита у растительноядных видов (например, у грызунов на его долю приходится до 53% от всей длины кишечника, а у хищных - до 22%). Особое значение для переработки целлюлозы имеет слепая кишка, поскольку животные не способны самостоятельно усваивать этот полисахарид. Однако микроорганизмы, населяющие слепую кишку, способны синтезировать нужные ферменты, следовательно, толстая кишка травоядных представляет собой своеобразное «микробиологическое производство», которое обеспечивает животное доступными соединениями.
Травоядные, у которых сбраживание происходит не в желудке, а в слепой кишке (например, зайцеобразные и многие грызуны), выделяют экскременты, в которых содержится много полезных веществ, поэтому животное обычно поедает этот помет. Когда помет повторно проходит через пищеварительный тракт, из помета всасываются нужные вещества, а неиспользованные шлаки удаляются во время дефекации. Коала выделяют в период кормления помет, богатый питательными веществами, которым кормят детеныша. Поедание экскрементов называется копрофагией.
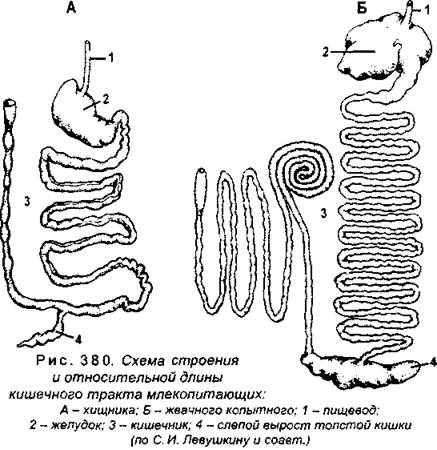
Толстая кишка заканчивается прямой кишкой, которая у млекопитающих хорошо выражена, она открывается наружу анальным отверстием. У однопроходных (утконоса, ехидны) кишечник открывается в клоаку. Общая длина кишечника у млекопитающих всегда больше длины тела, например, у хищника ласки в 2,5 раза, а у травоядной овцы в 29 раз.
Дыхательная система представлена дыхательными путями и респираторным отделом. Дыхательные (воздухоносные) пути проводят воздух, очищают его от пылевых частиц и микроорганизмов, согревают и увлажняют, в респираторном отделе происходит газообмен между кровью и легочным воздухом. Слизистая оболочка воздухоносных путей (кроме глотки и части гортани) выстлана многорядным реснитчатым эпителием с большим количеством бокаловидных клеток, которые выделяют слизь, покрывающую снаружи слизистую оболочку. Биение ресничек гонит слизистый слой с прилипшими к нему пылевыми частицами по направлению к глотке, после чего эта слизь проглатывается.
Верхние дыхательные пути включают в себя носовую полость и часть глотки (носоглотку и ротоглотку), нижние - гортань, трахею и систему бронхов разного калибра. В носовой полости, которая у млекопитающих отделена от ротовой твердым нёбом, имеется система носовых ходов, благодаря которым поверхность полости значительно увеличивается. Именно там, главным образом, воздух нагревается, очищается и увлажняется, кроме того, в носовой полости имеется обонятельный отдел, выстланный сенсорным эпителием.
Гортань млекопитающих имеет более сложное строение, чем у других классов позвоночных, она выполняет две функции - проведение воздуха и голосообразующую. Стенки гортани укреплены не только перстневидным и черпаловидными хрящами, но и характерным для млекопитающих крупным щитовидным хрящом, кроме того, у млекопитающих имеется эластический хрящ - надгортанник, прикрывающий вход в гортань во время глотания пищевого комка. В гортани расположены голосовые складки, в толще которых имеются эластические голосовые связки, над ними находятся ложные голосовые складки, не содержащие эластических волокон.
Полость гортани разделена на три отдела: верхний - преддверие гортани (пространство от входа в гортань до ложных голосовых складок); средний - желудочки гортани (пространство между ложными и истинными голосовыми складками) и нижний - подголосовая область (пространство, расположенное ниже голосовых складок, до нижнего края перстневидного хряща). Образование звука происходит при изменении натяжения голосовых связок и изменении расстояния между голосовыми складками (голосовой щели) за счет многочисленных мышц гортани (мышц, изменяющих натяжение голосовых складок, и мышц, изменяющих ширину голосовой щели). Остальные мышцы изменяют положение гортани как органа в целом (над- и подподъязычные мышцы) или изменяют положение надгортанника.
Стенки трахеи укреплены хрящевыми полукольцами, свободные концы которых закрыты мембраной, что весьма полезно, поскольку мембрана не мешает прохождению пищевого комка по расположенному позади трахеи пищеводу. Нижний конец трахеи делится на два бронха (главные, или бронхи первого порядка), которые проникают в ткань правого и левого легких и там многократно ветвятся на более мелкие бронхи, образуя в совокупности бронхиальное дерево. Мелкие бронхи называются бронхиолами. Стенки более крупных бронхов укреплены хрящами, но у самых мелких бронхиол хрящи отсутствуют, но снабжены более выраженной мускулатурой.
Морфофункциональной единицей легкого млекопитающих является легочный ацинус, который образуется в результате ветвления терминальной бронхиолы. При этом последовательно образуются респираторные бронхиолы I, II и III порядков, последние заканчиваются альвеолярными ходами, от которых отходят боковые выросты в виде мешочков с очень тонкими стенками - альвеолы. Пространство между ацинусами заполнено соединительной тканью - интерстицием. Газообмен в легких млекопитающих происходит в альвеолярных ходах и альвеолах, стенки которых густо оплетены кровеносными сосудами. Процесс газообмена подробно рассмотрен в разделе «Дыхательная система» человека (см. т. 1). Количество альвеол в легких млекопитающих очень велико, например, в легких ленивцев, которые отличаются своей малоподвижностью, имеется около 6 млн., а у подвижных хищных - около 300 - 500 млн.
Кроме газообмена, дыхательная система выполняет и другие функции, в частности, через легкие выделяются некоторые продукты обмена. Дыхательные пути участвуют в терморегуляции, например собак, кожа которых крайне бедна потовыми железами, при повышении температуры воздуха они начинают более глубоко дышать, при этом через широко раскрытую пасть активно испаряется слюна, охлаждая животное.
Дыхательные движения у млекопитающих осуществляются двумя способами: за счет изменения объема грудной клетки (реберное дыхание) и за счет деятельности диафрагмальной мышцы (диафрагмальное дыхание). Частота дыхательных актов у разных видов неодинакова, зависит от многих показателей: размера и состояния животного, физической нагрузки, образа жизни, внешней температуры (табл. 9) и др. Водные млекопитающие (ластоногие, китообразные) способны длительное время находиться под водой, например, кашалот - свыше часа.
Кровеносная система млекопитающих, как и птиц, характеризуется полным разделением артериальной и венозной крови, поэтому все органы (кроме печени) снабжаются чистой артериальной кровью. Четырехкамерное сердце состоит из двух предсердий и двух желудочков, при этом межжелудочковая перегородка млекопитающих не гомологична частичной перегородке, которая имеется в желудочке пресмыкающихся, а развивается совершенно другим путем. Правая половина сердца, содержащая только венозную кровь, полностью изолирована от левой половины, в которой находится артериальная кровь. Венозный синус (пазуха) и артериальный конус в сердце млекопитающих отсутствуют. У млекопитающих два круга кровообращения: большой и малый, некоторые авторы выделяют и третий - сердечный круг.
Таблица 9
Частота дыхания в 1 мин. у разных видов млекопитающих в зависимости от температуры внешней среды (по Калабухову)
|
Виды животных |
Температура внешней среды (в °С) |
||||
|
+5-+10 |
+10-+15 |
+15-+20 |
+20 - +25 |
+25 - +30 |
|
|
Лисица |
26 |
26 |
29 |
39 |
— |
|
Медведь белый |
15 |
15 |
25 |
30 |
_ |
|
Медведь черный |
8 |
15 |
15 |
24 |
41 |
|
Марал |
17 |
18 |
39 |
— |
— |
|
Верблюд |
7 |
9 |
9 |
9 |
12 |
Артериальная система большого круга кровообращения начинается с отходящей от левого желудочка единственной системной дуги аорты, которой у млекопитающих является левая дуга (напомним, что у птиц, наоборот, сохраняется только правая системная дуга аорты). Дуга аорты огибает левый бронх, поворачивает вниз и проходит через все туловище. Часть аорты, которая проходит через грудную клетку, называется грудной аортой, через специальное отверстие в диафрагме она проникает в брюшную полость и проходит ее в виде брюшной аорты. На всем протяжении аорта отдает артерии разной толщины к стенке тела (париетальные ветви) и к внутренним органам (висцеральные ветви), в области таза брюшная аорта разделяется на две мощные общие подвздошные артерии, каудально продолжается тонкая хвостовая артерия. Общие подвздошные артерии делятся на внутренние (снабжают кровью органы таза) и наружные (кровоснабжают задние конечности).
От верхней (передней) части дуги аорты обычно отходят три крупных сосуда: безымянная артерия (плечеголовной ствол), левая общая сонная и левая подключичная. Часто левая общая сонная артерия отходит от безымянной артерии. Короткая безымянная артерия вскоре делится на правую общую сонную и правую подключичную артерии. Общая сонная артерия (правая и левая) вскоре делится на две ветви - наружную (кровоснабжает преимущественно органы лицевого черепа) и внутреннюю (кровоснабжает головной мозг и орган зрения) сонные артерии. Подключичные артерии, в основном, кровоснабжают передние конечности, а также шею и стенку грудной клетки.
Венозная система. Венозная кровь от головы оттекает по наружным и внутренним яремным венам, которые с каждой стороны тела сливаются с соответствующей подключичной веной. В результате образуются правая и левая передние полые вены. У некоторых видов обе передние полые вены самостоятельно впадают в правое предсердие, однако у большинства млекопитающих левая сливается с правой полой веной, которая и впадает в сердце. При этом левый кювьеров проток видоизменяется в коронарный синус, в который впадают собственные вены сердца, этот синус открывается непосредственно в правое предсердие.
Венозная кровь задней части тела в конечном итоге собирается в заднюю полую вену, которая образуется при слиянии хвостовой вены с подвздошными венами. По пути к сердцу задняя полая вена принимает в себя вены, идущие от внутренних органов, проходит через печень (но не участвует в образовании воротной системы печени), по выходу из нее принимает крупные печеночные вены, затем подходит к сердцу и впадает в его правое предсердие.
Воротная система почек у млекопитающих не образуется, зато у них присутствуют непарная и полуне- парная вены, которые являются рудиментами задних кардинальных вен. Непарная вена самостоятельно впадает в переднюю полую вену, а полунепарная (левая непарная) обычно впадает в непарную (правую) вену.
Воротную систему печени образует только одна очень мощная воротная вена печени, которая образуется при слиянии венозных сосудов, несущих кровь от кишечника, поджелудочной железы и селезенки.
Малый круг не имеет существенных особенностей. От правого желудочка отходит легочный ствол, который вскоре делится на две легочные артерии - правую и левую. Каждая из них входит в одноименное легкое и распадается на сеть капилляров, оплетающих стенки альвеол, где происходит газообмен. Насыщенная кислородом артериальная кровь собирается в легочные вены, по которым кровь поступает в левое предсердие.
Кровеносная система млекопитающих отличается высоким сердечным индексом (см. табл. 3), относительно большой частотой сердечных сокращений, быстрым кровотоком, большим объемом циркулирующей крови (см. табл. 4), высоким давлением крови в сосудах (например, у слона 120/90 мм рт. ст., у собаки - 112/56). Существенной особенностью крови млекопитающих является отсутствие ядер в зрелых эритроцитах, что весьма выгодно, поскольку позволяет заполнить объем клетки, который заполняло ядро, дополнительным количеством гемоглобина. В результате в 100 см3 крови млекопитающих содержится 10 - 15 г гемоглобина, тогда как у рыб и амфибий - всего 5 - 10 г. Все это, наряду с высокоэффективной системой газообмена в альвеолах легких, делает уровень обменных процессов очень высоким (у млекопитающих и птиц скорость метаболизма примерно в 20 раз выше, чем у современных рептилий). Поскольку окислительно-восстановительные реакции, осуществляющиеся в клетках, являются экзотермическими, они сопровождаются выделением большого количества тепла. Поэтому млекопитающие, наряду с птицами, являются гомойотермными животными.
Лимфатическая система развита хорошо, она состоит из лимфатических капилляров и сосудов, которые проходят через лимфатические узлы. Основная масса лимфы впадает в кровеносное сосудистое русло в области яремных вен.
Выделительная система млекопитающих представлена двумя компактными тазовыми почками, расположенными в брюшной полости. Поверхность почки обычно гладкая, но у некоторых видов (например, китообразных) почки дольчатые. Вещество почки ясно делится на корковое и мозговое, имеющие разную окраску. На переднем конце почки располагается надпочечник, который является железой внутренней секреции и к почке функционального отношения не имеет.
Морфофункциональной единицей почки млекопитающих является нефрон маммального типа, т.е. между проксимальным и дистальным извитыми канальцами имеется петля Генле, которая участвует в концентрировании мочи (рис. 381). Более подробно о строении и функциях нефрона человека, а также механизме образования первичной мочи говорится в разделе «Мочевые органы» (см. т. 1). Для почек млекопитающих характерно очень большое количество нефронов, например, в обеих почках мыши насчитывается около 10000 нефронов, а у кролика - 285000.
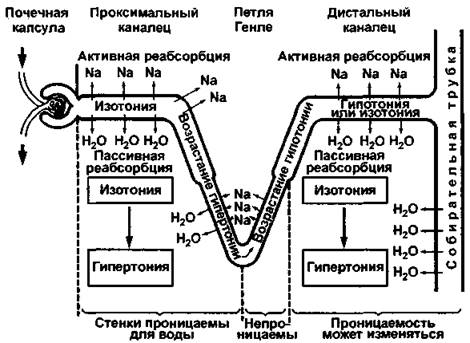
Рис. 381. Схема функционирования концентрационного аппарата в почке млекопитающих (по Ноар)
Продуктом азотистого обмена у млекопитающих, в основном (на 68 - 91%), является мочевина, которая более токсична, чем мочевая кислота, но так как моча имеет относительно большой объем (по сравнению с рептилиями или птицами), то негативных последствий это не несет. Напомним, что обычно выделяется не какой-либо один тип продуктов обмена, а несколько, но один из них является основным, например, млекопитающие также выделяют мочевую кислоту, но ее количество незначительно (0,1 - 8%).
Вторичная моча собирается в почечной лохани, от которой начинается мочеточник, впадающий в мочевой пузырь. Он расположен в малом тазу и служит накоплению мочи. Моча выводится через мочеиспускательный канал, строение которого у самцов и самок неодинаково. У самцов мочеиспускательный канал объединен с дистальным отделом половых путей и проходит через губчатое тело полового члена. У самок мочеиспускательный канал представляет собой самостоятельную короткую трубку, открывающуюся наружным отверстием мочеиспускательного канала в дистальный отдел влагалища - мочеполовой канал (видоизмененный мочеполовой синус). У однопроходных мочеточники впадают в мочеполовой синус, оттуда моча попадает в клоаку.
Кроме почек, выделительную функцию у млекопитающих выполняют легкие, кожа (потовые железы выделяют соли и мочевину) и толстая кишка. Однако таким образом выводится всего около 3% продуктов азотистого обмена.
Половая система. Мужская половая система состоит из парных семенников овальной формы, половых протоков, семенных пузырьков, предстательной железы и полового члена (рис. 382-А). Семенники у всех млекопитающих закладываются в брюшной полости, но только у однопроходных, отдельных неполнозубых и насекомоядных, слонов и китообразных они остаются там и у взрослых особей. У самцов других групп они через паховый канал выходят из брюшной полости, предварительно все слои брюшной стенки вытягиваются, образуя особый мешочек - мошонку, которая имеется только у млекопитающих (рис. 383). В мошонке находятся не только семенники, но также их придатки (как и у других амниот, они образуются из канальцев переднего отдела мезонефроса), начальная часть семяпровода, сосуды и нервы.
Половые клетки, образующиеся в стенках извитых семенных канальцев семенника (яичка), поступают в канальцы придатка, где они обретают подвижность и скапливаются до эякуляции. От придатка начинается семяпровод, который происходит из вольфова протока. Семяпроводы имеют значительную длину, поскольку они возвращаются в брюшную полость и в области таза впадают в мочеиспускательный канал, предварительно образовав боковые ответвления - семенные пузырьки, секрет которых входит в состав спермы.
Мочеиспускательный канал самцов млекопитающих проходит через губчатое тело полового члена, у основания которого располагается предстательная железа - железисто-мышечный орган, продуцирующий компоненты спермы и множество биологически активных веществ. В момент наивысшего полового возбуждения в ходе полового акта сокращение гладких мышечных клеток проталкивает сперму, благодаря чему она с силой выбрасывается из полового члена во влагалище самки.
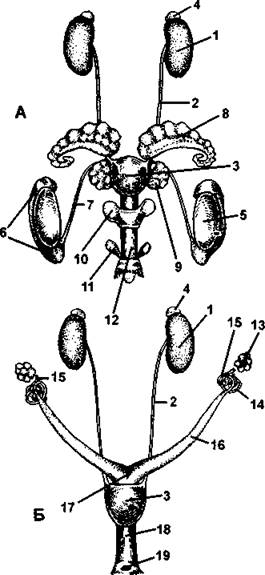
Рис. 382. Мочеполовая система крысы:
А - самец; Б - самка; 1 - почка; 2 – мочеточник; 3 - мочевой пузырь; 4 – надпочечник; 5 – семенник; 6 - придаток семенника; 7 – семяпровод; 8 - семенной пузырек; 9 - предстательная железа; 10 - куперова железа; 11 - препуциальнаяжелеза; 72 - половой член; 13 - яичник; 14- яйцевод; 15- воронка яйцевода; 16- рог матки; 17 - матка; 18- влагалище; 19 - половое отверстие
Половой член млекопитающих организован значительно сложнее, чем копулятивный орган крокодилов и черепах. Он образован двумя пещеристыми телами, которые продольно срастаются друг с другом и с губчатым телом, заканчивающимся головкой, окруженной кожной складкой - крайней плотью. У многих млекопитающих (хищных, ластоногих и др.) в соединительной ткани между пещеристыми телами располагается еще особая длинная кость бакулюм (os penis), которая упрочняет половой член и в значительной мере увеличивает надежность эрекции. Эта кость может быть весьма массивной, например, умельцы некоторых народов Севера из соответствующих костей ластоногих изготовляют различные предметы (вплоть до этажерок!), и не случайно именно пенис моржа давно стал в народе красноречивой метафорой, подчеркивающей физическую незаурядность мужских достоинств. Мочеиспускательный канал проходит через губчатое тело и открывается наружным мочеиспускательным отверстием. В отличие от рептилий, совокупительный орган которых лежит внутри клоаки, значительная часть пениса млекопитающих представляет собой наружный орган.
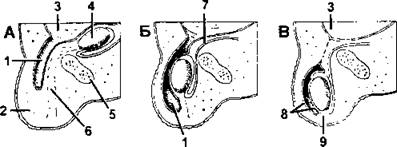
Рис. 383. Опускание семенника у млекопитающих; брюшная поверхность тела обращена влево.
Из полости тела выпячивается влагалищный отросток (А); выстилающая его брюшина формирует влагалищную оболочку (tunica vaginalis) кармана мошонки (Б). На В штриховой линией обозначено положение пахового канала у тех млекопитающих, у которых карман мошонки не замыкается полностью: 1 - влагалищный отросток; 2 - мошонка; 3 - брюшная полость; 4 - семенник; 5 - симфиз; 6 - направляющая связка; 7 - семяпровод; 8 - влагалищная оболочка; 9 - направляющая связка (по Тюрнеру)
Строение полового члена однопроходных во многом напоминает совокупительный орган самцов крокодилов и черепах (рис. 384).
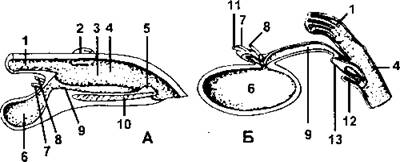
Рис. 384. Разрез клоаки:
А - самца черепахи; Б - самца однопроходного млекопитающего - ехидны. У черепахи в дне клоаки располагается образование, подобное пенису: парные гребни способны временно смыкаться с образованием трубки для выведения спермы. У однопроходных внутри клоаки сформирован пенис; внутри него проходит разветвленный канал для выведения спермы, но моча выходит наружу мимо него через клоаку. У большинства рептилий мочеточник открывается в клоаку сверху, далеко от мочевого пузыря; по положению его выхода черепахи похожи на млекопитающих в большей степени - у них он переместился вентрально. Желоб на А и трубка на Б, обозначенные как мочеиспускательный канал, по существу, являются лишь вентральным передним отделением клоаки. К особенностям черепах относится присутствие на боковых стенках клоаки пары маленьких «дополнительных» образований, подобных мочевому пузырю; 1 - кишка; 2 - анальный мешок; 3 - одна из парных складок - пещеристых тел члена; 4 - клоака; 5 - головка полового члена; 6 - мочевой пузырь; 7 - семяпровод; 8 - мочеточник; 9 - мочеиспускательный канал; 10 - волокнистая ткань; 11 - рудиментарный яйцевод; 12 - половой член в сумке; 13 - канал полового члена для спермы (из А. Ромера и соавт.)
Женская половая система представлена парными яичниками и половыми путями (см. рис. 382-Б). Яичники всегда находятся в брюшной полости, где фиксированы на задней (спинной) стенке брыжейкой. Яйцеводы, возникшие из мюллеровых протоков, непосредственно не связаны с яичниками (как и у всех других позвоночных), они открываются в брюшную полость отверстиями воронок, расположенными вблизи яичников.
Яйцеводы млекопитающих лучше других позвоночных дифференцированы на отделы. Первый отдел, заканчивающийся воронкой, носит названия маточной (фаллопиевой) трубы. Далее следует более широкий отдел - матка, которая с каждой стороны тела впадает в непарное влагалище, проксимальный отдел которого образуется из мюл- лерова протока, а дистальный - мочеполовой канал - из мочеполового синуса (пазухи). В дистальный отдел (при большой длине он выглядит как продолжение влагалища) также открывается наружное отверстие женского мочеиспускательного канала. Там же находится гомолог пениса - клитор, который устроен проще: он состоит из двух пещеристых тел и головки, губчатого тела нет.
У разных систематических групп млекопитающих строение женских половых органов может существенно различаться. У самых примитивных млекопитающих - однопроходных (утконос, ехидна, проехидна) влагалища нет, яйцеводы, раздельные на всем протяжении, дифференцируются на маточные трубы и матки, которые самостоятельно открываются в мочеполовой синус. У сумчатых влагалище имеется, но оно часто бывает парным, представляя собой пару трубок, сросшихся своими проксимальными и дистальными (т.е. передними и задними) отделами (рис. 385).
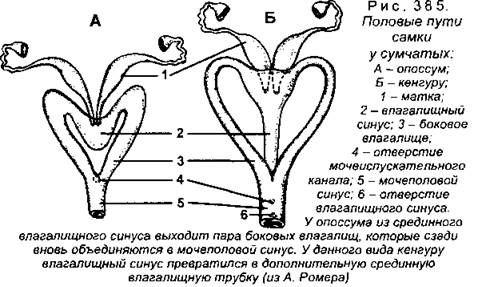
Влагалище плацентарных млекопитающих всегда непарное, но проксимальные отделы в той или иной степени сохраняют парность (рис. 386). У многих неполнозубых, грызунов и летучих мышей матка двойная, при этом правая и левая матка полностью обособлены одна от другой и открываются во влагалище самостоятельными отверстиями. Если матки соединяются друг с другом своими дистальными отделами, то матка называется двураздельной, она имеется у некоторых хищных, грызунов и летучих мышей. Матки могут срастаться своими проксимальными отделами, при этом с маточными трубами связаны лишь самые проксимальные части (в виде рогов) - это двурогая матка, она имеется у некоторых хищных, копытных и китообразных. У высших млекопитающих (приматов) и некоторых летучих мышей матка целиком непарная и по бокам соединена с маточными трубами (которые все равно остаются парными!), такая матка называется простой.
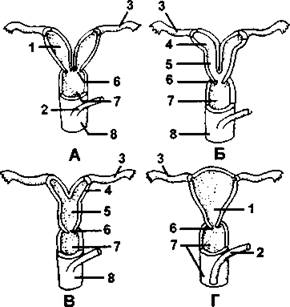
Рис. 386. Схемы, показывающие постепенное объединение задних частей яйцеводов (маток) у плацентарных млекопитающих. Матка и часть влагалища вскрыты:
А - двойная матка; В - двураздельная матка; В - двурогая матка; Г - простая матка; 1 - матка; 2 - мочеиспускательный канал; 3 - фаллопиева труба; 4 - рог матки; 5 - тело матки; 6 - шейка матки; 7 - влагалище; 8 - мочеполовой синус (из А. Ромера и соавт.)
Развитие. Оплодотворение, как и у всех амниот, внутреннее. Сперма попадает в половые пути самки во время полового акта, в ходе которого затвердевший эрегированный половой член самца вводится во влагалище самки. Оплодотворение яйцеклетки и начальные этапы развития проходят в маточных трубах. Однако дальнейшее развитие млекопитающих, принадлежащих к разным систематическим группам, идет неодинаково. Яйца млекопитающих содержат мало желтка (изолецитальные яйца), дробление асинхронное, т.е. бластомеры делятся не одновременно, поэтому увеличение числа бластомеров зародыша происходит не в геометрической прогрессии (2 → 4 → 8 → 16 → 32 → 64 и т.д.), а постепенно, в результате количество бластомеров в зародыше может быть нечетным. Кроме того, для раннего развития зародыша млекопитающих характерна компактизация, в ходе которой бластомеры внезапно сближаются и в большей степени контактируют друг с другом (рис. 387).
Наиболее примитивные - однопроходные - откладывают яйца, покрытые скорлупой, однако, в отличие от рептилий и птиц, яйцо содержит не зиготу и питательные вещества, а зародыш, находящийся на средней стадии своего развития. Первую половину своего развития зародыш прошел в половых путях матери.
Рис. 387. 8-клеточный зародыш мыши: до (А) и после (Б) компактизации; микрофотографии получены с помощью сканирующего электронного микроскопа (из С. Гилберта)
Достигнув определенного состояния, зародыш разрушает изнутри скорлупу яйца и выходит из него, при этом у ехидн он оказывается в сумке, куда самка помещает яйцо, а у утконосов - в гнезде (у них нет сумки). Как и все млекопитающие, однопроходные выкармливают детенышей молоком, которое у них стекает прямо по шерсти, поскольку протоки млечных желез не образуют соска.
У сумчатых развивающийся зародыш контактирует в области желточного мешка с гипертрофированной стенкой матки и диффузно осуществляет с ней обмен веществ. Однако полного развития в половых путях у сумчатых не происходит, поэтому рождается еще, по сути, эмбрион очень маленького размера (например, у гигантского рыжего кенгуру, рост которого превышает 2 м, рождается эмбрион длиной всего около 3 см, который немедленно ползет в сумку (причем самостоятельно!), обхватывает сосок и держит его очень прочно (по этой причине некоторое время даже считали, что кенгуру размножаются почкованием). Питаясь молоком матери, детеныш быстро заканчивает свое развитие, но еще довольно долго остается в сумке.
Развитие высших млекопитающих - плацентарных - целиком проходит в женских половых путях. Оплодотворенная в маточной трубе яйцеклетка (зигота) там же приступает к дроблению, одновременно перемещаясь в полость матки. Там наружный слой зародыша - трофобласт, на поверхности которого образуются ворсинки хориона, - срастается со стенкой матки, слизистая оболочка которой к тому моменту сильно утолщена. Со временем между развивающимся зародышем и стенкой матки образуется общая связующая структура - плацента, через которую происходит обмен веществ между матерью и зародышем. По завершении внутриматочного развития плацентарные млекопитающие рождают хорошо развитого детеныша и некоторое время вскармливают его молоком.
Подобно птенцам птиц, которых, напомним, делят на выводковые и гнездовые, детеныши млекопитающих могут рождаться зрячими, подвижными и покрытыми шерстью, например жвачные, или слепыми, голыми и совершенно беззащитными, например хищные. Отличия могут быть даже среди близких видов, например, зайчата рождаются зрячими и вскоре после рождения могут бегать, тогда как крольчата слепые, голые и беспомощные.
Млекопитающие проявляют хорошо выраженную заботу о потомстве, при этом детеныши выкармливаются сначала молоком, потом пищей взрослых, причем хищные специально для этого приносят часть пойманной добычи. Родители охраняют детенышей и вместе с другими взрослыми особями обучают. Однако через определенное время молодые самцы чаще всего изгоняются, чтобы предотвратить кровосмешение.
Нервная система. Центральная нервная система млекопитающих является наиболее сложной среди всех позвоночных. Головной мозг очень крупный и по массе значительно превосходит спинной мозг (рис. 388). Так, например, у хищных головной мозг соотносится со спинным в пропорции 4:1, у приматов -8:1, у человекообразных обезьян - 20-25:1, а у человека - 45:1. Особенно крупным является конечный мозг, по данным Никитенко, масса полушарий конечного мозга у ежей составляет 48% от массы всего головного мозга, у белок - 53%, у волков - 70%, у дельфинов - 75%. При этом увеличение достигается путем масштабного разрастания свода, а не полосатых тел, как это происходит у птиц. Свод у млекопитающих вторичный (нео- паллиум), поскольку вся его поверхность покрыта корой, образованной серым веществом. Кора полушарий конечного мозга (большого мозга) млекопитающих подразделяется на древнюю кору, которая имеет наиболее примитивное однослойное строение, похожее на участки коры полушарий рептилий и птиц, старую кору, которая также однослойная, но отделена от нижележащей подкорки корковой пластинкой, и новую кору, которая имеет наиболее сложное строение и состоит из нескольких слоев (у человека до шести в разных участках). Соотношение перечисленных участков у разных видов млекопитающих неодинаково и зависит прежде всего от уровня сложности поведения. Наиболее развита новая кора у хищных, приматов и особенно у человека, у которого на долю новой коры приходится около 96% всей коры, при этом новая кора отделена от старой и древней коры зонами средней (промежуточной) коры (перипалиокортикальной и периархикортикальной соответственно).

Рис. 388. Мозг кролика:
А – сверху; Б – снизу; В – сбоку; Г - продольный разрез; 1 - большие полушария; 2 - обонятельные доли; 3 - зрительный нерв; 4 - эпифиз; 5 - средний мозг; 6 - мозжечок; 7 - продолговатый мозг; 8 - гипофиз; 9 - варолиев мост; 10- мозговая воронка; 11 - мозолистое тело (по С. П. Наумову)
В коре полушарий большого мозга располагаются центры высшей нервной деятельности, они обеспечивают наиболее сложные поведенческие реакции, которые не заложены изначально, а развиваются в течение жизни, максимально эффективно приспосабливая животное к быстро меняющимся условиям внешней среды. Напомним, что безусловные поведенческие рефлексы (инстинкты), в том числе самые сложные, представляют собой определенную программу действий, за пределы которой животное не может выйти, даже если это необходимо. Кроме того, в коре располагаются высшие центры анализаторов. Таким образом, у млекопитающих кора конечного мозга становится ведущим отделом головного мозга, который интегрирует деятельность ЦНС. Мозг такого типа называют маммальным.
Усложнение поведения и развитие многочисленных условных рефлексов требует весьма значительной площади коры, прежде всего новой. У наименее подвижных млекопитающих (например, ленивцев) поверхность полушарий почти гладкая, но у большинства видов разросшаяся кора образует более или менее выраженные складки (извилины), разделенные бороздами, наиболее крупные' борозды отделяют доли полушарий, причем положение складок и борозд является характерным признаком для того или иного вида. В самом простом варианте имеется одна латеральная (сильвиева) борозда, которая отделяет височную долю, в более сложном головном мозге также центральная (роландова) борозда, отделяющая сзади лобную долю. У высших млекопитающих количество извилин и борозд может быть значительным, что существенно увеличивает общую поверхность коры. Особенно велика новая кора в области лобных долей, которые хорошо развиты у хищных и в еще большей степени у приматов. Именно там локализуются молодые ассоциативные центры, управляющие сложным поведением (высшая нервная деятельность).
В коре полушарий большого мозга выделяют проекционные поля, которые получают информацию от периферических (рецепторных) отделов анализаторов, между ними находятся ассоциативные поля, которые не связаны непосредственно с органами чувств, но образуют связи между проекционными полями, что делает аналитико-синтетическую обработку сигналов более качественной. Для коры характерно наличие сложнейших связей, которые функционально подразделяются на три группы: проекционные - связывают головной мозг с другими отделами ЦНС, при этом они могут быть восходящими (идут от нижележащих отделов к коре) или нисходящими (идут к нижележащим отделам); комиссуральные - связывают участки серого вещества правого и левого полушарий (правое и левое полушария связывает мощная перемычка из нервных волокон - мозолистое тело); ассоциативные - соединяют между собой нервные центры, расположенные в одном полушарии.
Базальные ядра (ядра полосатого тела и некоторые другие), которые у рептилий и птиц интегрируют деятельность всего головного мозга, у млекопитающих подчинены коре и выполняют роль подкорковых центров, контролирующих движения и мышечный тонус. Обонятельные луковицы млекопитающих развиты хорошо, особенно у тех видов, для которых обоняние является ведущим органом чувств.
Промежуточный мозг включает в себя зрительные бугры (таламусы), представляющие собой крупные подкорковые скопления серого вещества, надбугровую область (эпифиз, подвешенный к таламусам на поводках), забугровую область (она связана с органами зрения и слуха) и подбугровую (гипоталамус, связанный воронкой с гипофизом). Гипоталамус имеет связи не только с нейрогипофизом (задней долей), но и с аденогипофизом (передней долей) посредством сосудистой сети воротной системы гипофиза, куда выделяются рилизинг- факторы, синтезируемые нейросекреторными клетками гипоталамуса, которые регулируют синтез и поступление в кровь гормонов аденогипофиза. Средняя доля гипофиза малофункциональна. Промежуточный мозг сверху не виден, потому что прикрыт огромными полушариями конечного мозга.
В крыше среднего мозга обособляется характерная для млекопитающих пластинка четверохолмия, состоящая из двух бугорков, из которых верхняя пара связана со зрительным, а нижняя - со слуховым анализаторами.
Напомним, что у всех прочих классов позвоночных крыша среднего мозга разделена продольной бороздой на два бугра (двухолмие). Проводящие пути локализованы в ножкам среднего мозга. Размеры этого отдела головного мозга у млекопитающих невелики, что связано с его подчиненностью высшим центрам, прежде всего коре. От среднего мозга отходят III и IV пары черепных нервов. Полость среднего мозга представлена узким сильвиевым водопроводом, соединяющим между собой третий и четвертый желудочки.
Ромбовидный мозг млекопитающих разделен на задний и продолговатый. Задний мозг представлен не только мозжечком, но и хорошо выраженным мостом, что связано с более высокой организацией проводящих путей у млекопитающих, значительная их часть проходит через мост.
Мозжечок развит очень хорошо, состоит из средней части - непарного червя и боковых отделов - полушарий, также имеются клочок и узелок. Таким образом, наряду с древними (клочок, узелок) и старыми (передняя часть червя) структурами, у млекопитающих имеются более молодые (полушария и задняя часть червя). При этом более старые структуры получают чувствительную информацию от органа равновесия и спинного мозга (вестибуло- и спинно-мозжечковые пути), тогда как молодые - от коры полушарий конечного мозга (корково-мостомозжечковые пути). Наряду с полушариями большого мозга, мозжечок снаружи покрыт корой (правда, более простого строения, чем полушария большого мозга) серого вещества, именно в ней локализуются наиболее молодые центры, управляющие координацией сложных движений. На поверхности полушарий мозжечка, так же как и на поверхности полушарий конечного мозга, образуются складки - листики мозжечка, которые увеличивают поверхность коры. Более простыми функциями управляют ядра, расположенные в мозжечке медиально.
Столь сложная деятельность мозжечка млекопитающих нуждается в совершенной и многосторонней связи с ведущими отделами головного мозга, эти связи локализуются в ножках мозжечка (верхних, средних и нижних), при этом верхние (или передние) ножки направляются к среднему мозгу (они несут главным образом двигательные волокна), средние (несут импульсы из коры полушарий большого мозга) — к мосту, а нижние (или задние) - к продолговатому мозгу (несут проприоцептивные импульсы от спинного мозга).
В продолговатом мозге различают парные валики белого вещества - пирамиды, расположенные на передней поверхности продолговатого мозга. Пирамиды представляют собой двигательные пути, которые направляются от коры большого мозга к ядрам передних рогов спинного мозга и управляют деятельностью скелетной мускулатуры, именно эти проводящие пути обеспечивают произвольные движения.
В передней части спинного мозга и стволе головного мозга серое вещество находится не только в составе выраженных клеточных скоплений (ядер), но также диффузно распределено в виде ретикулярной формации. Ее значение у млекопитающих не столь велико, как у низших позвоночных, однако она участвует в координации простых движений, кроме того, ретикулярная формация принимает участие в передаче двигательных импульсов от вышележащих отделов мозга к нижележащим. Ретикулярная формация также стимулирует деятельность высших центров коры полушарий большого мозга и контролирует некоторые рефлексы (например, кашель, чихание).
Спинной мозг имеет характерное сегментарное строение, в переднем и заднем отделах образует утолщения (шейное и пояснично-крестцовое, соответственно). У взрослых особей длина спинного мозга меньше длины позвоночного столба, поэтому в задней части позвоночного канала находятся корешки спинного мозга, образующие конский хвост. Задняя, редуцированная, часть спинного мозга образует соединительнотканную концевую нить.
Периферическая нервная система. От головного мозга отходят 12 пар черепных нервов, среди которых I, II, и VIII пары являются чувствительными; IV, VI, XI и XII двигательными; III, V, VII, IX и X смешанными, причем III пара содержит двигательные и парасимпатические волокна, V - чувствительные и двигательные, VII, IX и X - чувствительные, двигательные и парасимпатические волокна. При этом у млекопитающих XI пара (добавочный нерв) полностью дифференцирована. Спинномозговые нервы (передние ветви) образуют нервные сплетения.
Вегетативная нервная система млекопитающих развита очень хорошо. Центры парасимпатического отдела находятся в вегетативных ядрах соответствующих черепных нервов (они содержат только парасимпатические волокна), а также в крестцовом отделе спинного мозга. Периферический отдел включает в себя упомянутые волокна черепных нервов вместе с промежуточными ганглиями, где белые соединительные волокна переключаются на серые (подробнее об этом см. т. 1 «Вегетативная (автономная) нервная система»). Центры симпатического отдела находятся в боковых рогах серого вещества некоторых сегментов спинного мозга (грудных, а также последнем шейном и верхних поясничных). Периферический отдел - это симпатические стволы, симпатические ганглии и сплетения.
Органы чувств у большинства млекопитающих развиты очень хорошо, частичная или полная редукция того или иного анализатора обычно носит вторичный характер и связана с особенностями образа жизни.
Орган зрения имеется у большинства млекопитающих. Строение глаза типично для наземных позвоночных (выпуклая роговица и двояковыпуклый хрусталик), аккомодация менее совершенна, чем у птиц, осуществляется только путем изменения кривизны хрусталика под воздействием ресничной мышцы. У мелких грызунов (например, мыши) аккомодация вообще отсутствует.
Различать цвета могут далеко не все млекопитающие, например, лесной хорек и многие другие животные лишены цветового зрения, другие виды различают лишь отдельные цвета спектра, и только очень немногие млекопитающие (высшие приматы восточного полушария) имеют полноценное цветовое зрение. Большинство видов способно различать только движущиеся предметы, оставляя неподвижные без внимания. Это связано с тем, что у млекопитающих преимущественно развиты другие органы чувств, а зрение для ориентации играет значительно меньшую роль, чем у птиц. Более острое зрение имеют обитатели степей, саванн, прерий и других обширных открытых территорий, а также ночные животные. Напротив, лесные млекопитающие видят хуже. У некоторых видов, преимущественно подземных (слепыша, некоторых кротов) зрение вообще атрофировалось, а редуцированные глаза полностью закрыты перепонкой.
Орган слуха у млекопитающих развит очень хорошо, а его строение гораздо более сложно, чем у других классов. Только у млекопитающих имеется наружное ухо, представляющее собой воронку из эластического хряща, покрытую кожей. Благодаря наличию хорошо развитых ушных мышц (они относятся к мимическим) большинство зверей способны поворачивать ушные раковины по направлению к источнику звука, четко определяя его местонахождение. Это позволяет безошибочно ориентироваться даже без участия зрения. В барабанной полости среднего уха находятся не одна, как у всех других наземных позвоночных, а три слуховые косточки - молоточек, наковальня и стремечко (об их развитии мы говорили выше, обсуждая череп), подвижно связанные между собой суставами, поэтому звукопередающий аппарат у млекопитающих совершеннее. Во внутреннем ухе имеется полностью сформированная улитка (рис. 389), в которой хорошо развит звуковоспринимающий кортиев орган. Особенно хорошо орган слуха развит у ночных видов и хищных, часто при этом слух является основным органом чувств (например, у кошачьих).
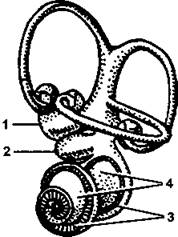
Рис. 389. Лабиринт млекопитающего: 1 - утрикулюс; 2 - саккулюс; 3 - основная мембрана; 4 - улитка
Часть млекопитающих способна ориентироваться с помощью эхолокации. В наибольшей степени это свойственно летучим мышам, землеройкам, ластоногим и дельфинам. При этом острота эхолокации очень высока, например, летучие мыши активно летают в темное время суток и неосвещенных пещерах, при этом уклоняясь от столкновений с предметами и разыскивая пищу, а дельфины, испуская звуки частотой 120 - 200 кГц, с высокой точностью определяют местонахождение косяков рыб на расстоянии до 3 км.
Орган обоняния развит исключительно хорошо, поскольку для большинства млекопитающих является основным в процессе ориентации. Обонятельная капсула имеет крупные размеры, область обонятельного эпителия весьма обширна. У ряда млекопитающих (сумчатых, грызунов, копытных) в обонятельной капсуле обособляется отдел, открывающийся в нёбно-носовой канал, это якобсонов орган (см. раздел «Пресмыкающиеся»). Обоняние вторично редуцированно у китов, однако у других морских млекопитающих (тюленей) обоняние развито очень хорошо. Острое обоняние помогает млекопитающим свободно ориентироваться по запахам, оставленным другими животными, поэтому запаховая сигнализация чрезвычайно широко распространена в природе. Чуткость обоняния при этом может быть удивительно высокой, например, свиньи и собаки легко отыскивают предметы даже под землей, поэтому их часто используют для поиска очень дорогих грибов - трюфелей.
Поведение млекопитающих наиболее сложное среди всех животных, оно основывается не на врожденных (безусловных) рефлексах, определяющих инстинктивную деятельность, а на приобретенных в течение жизни условных рефлексах. Для многих видов свойственно обучение молодняка, в ходе которого молодые особи получают сведения, которые накапливались в популяции в течение многих поколений. Например, слоны, ведомые вожаком - самой опытной слонихой, узнают место расположения всех жизненно важных мест, которые могут быть удалены друг от друга на большие расстояния, и в последующем находят дорогу самостоятельно. У птиц вся информация о сезонных миграциях заложена на уровне безусловных рефлексов.
Высокое развитие высшей нервной деятельности, основанной на условных рефлексах, формируемых в коре больших полушарий конечного мозга, позволяет млекопитающим проявлять великолепную экологическую пластичность. Поэтому млекопитающие сумели занять самые разнообразные экологические ниши и в настоящее время являются доминирующим классом среди позвоночных.
Анатомические критерии для определения степени психического развития организма не являются абсолютно надежными. В частности, абсолютная и относительная масса головного мозга не всегда существенна. Так, например, масса головного мозга макака составляет 62 г, гиббона - 100, шимпанзе и орангутана - 400 - 420, гориллы - 500, кошки - 30, дельфина - 1800, кита - 7000, слона - 5000 г и т.д.
По относительной массе мозга по сравнению с массой тела многие млекопитающие (ряд низших обезьян, грызунов и др.) и некоторые птицы превосходят человека. Российский антрополог Я. Я. Рогинский предложил оригинальный «квадратный указатель мозга» — произведение абсолютной массы мозга на относительную. Приведенный указатель отражает уровень цефализации, или «церебролизации», т.е. «величину массы мозга при исключенном влиянии массы тела на массу мозга» (Я.Я. Рогинский, М.Г. Левин). По данным авторов, эта величина составляет у насекомоядных 0,06; грызунов - 0,19; неполнозубых - 0,25; копытных и хищных - 1,14; полуобезьян и американских когтистых обезьян - 1,37; низших обезьян Старого Света - 2,25; гиббонов - 2,51; ластоногих - 2,81; китообразных - 5,25; человекообразных обезьян - 7,35; слонов - 9,82; человека - 32,0. Сравнительный анализ «квадратного указателя мозга» позволяет говорить «о связи интеллекта с деятельностью анализирующей конечности (цепкого хвоста, хобота, руки) у животных с большой массой мозга, уже не говоря про человека» (Я. Я. Рогинский).
Вопросы для самоконтроля и повторения
1. Какие признаки присущи хрящевым рыбам?
2. Опишите внешнее строение акулы.
3. Что собой представляет плавник?
4. Какое строение имеют покровы хрящевых рыб? Что такое плакоидная чешуя?
5. На какие отделы подразделяется скелет позвоночных животных?
6. Какие отделы выделяют в осевом скелете?
7. На какие отделы подразделяется череп?
8. Что такое добавочный скелет?
9. Каково строение мышечной системы рыб?
10. Как организована пищеварительная система позвоночных? Какие функции выполняют печень и поджелудочная железа?
11. Как происходит газообмен у хрящевых рыб?
12. Какое строение имеет кровеносная система рыб?
13. Какое строение имеет туловищная почка? Как она функционирует?
14. Какое строение имеет чешуя костных рыб?
15. Каковы особенности скелета костных рыб?
16. Какие функции выполняют электрические органы рыб?
17. Какое строение имеет жаберный аппарат костных рыб?
18. Опишите особенности газообмена костных рыб. Какие дополнительные органы газообмена имеются у костных рыб?
19. Как организована половая система хрящевых рыб? Костных рыб?
20. Как происходит развитие рыб? Приведите примеры.
21. Как организована центральная нервная система рыб? Какое строение имеет головной мозг?
22. Какое строение имеет орган зрения рыб?
23. Перечислите особенности строения наземных животных по сравнению с рыбами.
24. Какое строение имеют покровы тела земноводных? Почему они всегда влажные?
25. На какие отделы подразделяется позвоночный столб амфибий?
26. Каково происхождение подъязычной дуги наземных животных?
27. Чем характеризуется пятипалая конечность?
28. Как организована дыхательная система амфибий?
29. Чем вызвана необходимость второго круга кровообращения?
30. Каковы особенности кожных покровов рептилий?
31. Как дифференцирован позвоночный столб рептилий? Какие функции выполняют первые два шейных позвонка?
32. Чем характеризуется череп рептилий?
33. Как осуществляется дыхание у рептилий?
34. Опишите строение и функции сердца рептилий.
35. Каково строение тазовой почки? Как она функционирует?
36. Какой отдел головного мозга рептилий является ведущим?
37. Какие виды перьев птиц вы знаете? Каково их строение и функции? Как происходит развитие пера?
38. Какие особенности имеет скелет птиц? Чем они вызваны?
39. Как организована мышечная система птиц?
40. Как осуществляется газообмен у птиц?
41. Как происходит голосообразование у птиц?
42. Опишите кровеносную систему птиц.
43. Чем вызвана редукция правых половых путей самок птиц?
44. Как происходит размножение птиц?
45. Какое строение имеют кожные покровы млекопитающих? Какие производные эпидермиса вы знаете?
46. Каково значение кожи в терморегуляции у млекопитающих?
47. В связи с чем птицы и млекопитающие являются теплокровными животными? Какие преимущества дает постоянная температура тела? Как она поддерживается?
48. Какое строение имеют зубы млекопитающих? Что такое зубная формула? Как ее составляют?
49. Какое строение имеет пищеварительная система у млекопитающих, питающихся различной пищей?
50. Какие особенности имеет нервная система млекопитающих? Чем объясняется их сложное поведение?