Рептилии, птицы и млекопитающие образуют группу высших позвоночных - амниот, которые способны размножаться без внешних источников воды благодаря наличию амниотической полости, ограниченной зародышевой амниотической оболочкой. Соответственно, рыбы и земноводные составляют группу низших позвоночных - анамний, у которых такая оболочка отсутствует, поэтому у них размножение осуществляется в водоемах, т.к. оболочки яйца у них не способны защитить развивающийся эмбрион от потери воды на суше. Именно поэтому ареалы анамний, как правило, ограничены водоемами или прибрежными территориями. Наличие амниотической оболочки позволило амниотам не зависеть от водоемов в период размножения, поэтому они расселены гораздо более широко и занимают самые разнообразные экологические ниши.
Среди амниот наиболее примитивными являются пресмыкающиеся - единственные холоднокровные амниоты. В настоящее время насчитывается около 6000 видов рептилий, которые составляют четыре отряда: клювоголовые, чешуйчатые, крокодилы и черепахи. Огромное количество видов пресмыкающихся вымерло в меловом периоде мезозойской эры по причинам, которые до сих пор не вполне ясны. Большинство современных рептилий обитают на суше, в том числе в местах с очень сухим климатом (пустынях). Некоторые виды вторично перешли к водному образу жизни.
Внешнее строение рептилий достаточно разнообразно, но в типичных случаях тело подразделяется на голову, туловище и хвост. Тело черепах заключено в массивный костный панцирь, надежно защищающий тело животного.
Анатомия. Кожные покровы рептилий значительно более толстые, чем у амфибий, и, в отличие от них, сухие (рис. 310). Наружный слой представлен эпидермисом (многослойный ороговевающий эпителий), который у многих видов формирует утолщения - чешуи.
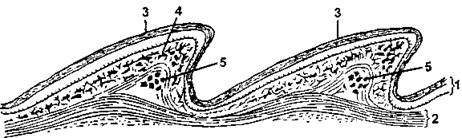
Рис. 310. Продольный разрез через кожу ящерицы (Lacerta):
1 - эпидермис; 2 - собственно кожа; 3 - роговой слой; 4 - пигментные клетки; 5 - кожные окостенения (по Бючли)
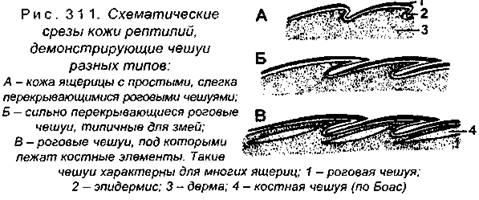
У многих ящериц роговые чешуи лишь слегка налегают друг на друга (рис. 311), при этом эпидермис между чешуями более тонкий, чем сами чешуи. У змей чешуи налегают друг на друга черепицеобразно, наконец у некоторых ящериц под чешуями лежат костные образования. Форма чешуй отличается не только у разных видов, но и зависит от расположения на теле животного. Обращаем внимание читателя на то, что костные чешуи рыб образуются из кориума, т.е. имеют мезодермальное происхождение, тогда как роговые чешуи пресмыкающихся происходят из эктодермального эпидермиса. Производными эпидермиса также являются когти, которыми вооружены пальцы.
Самый поверхностный роговой слой эпидермиса постоянно слущивается, пополнение клеточного состава осуществляют стволовые клетки, которые расположены в прилегающем к кориуму ростковом (мальпигиевом) слое. По мере деления этих клеток другие клетки, находящиеся над ними, оттесняются кнаружи, постепенно заполняются кератогиалином, отмирают и отслаиваются.
У змей периодически происходит линька, в процессе которой старый эпидермис сходит с тела наподобие чулка, при этом у гремучих змей на конце хвоста остается небольшой участок старой кожи, образующий характерную для них погремушку (рис. 312).

Сухая кожа рептилий содержит очень мало желез, у ящериц имеются железы, открывающиеся бедренными порами, у крокодилов железы имеются на спине, вблизи клоаки и на голове под нижней челюстью, более хорошо железы развиты у некоторых черепах. Под эпидермисом располагается кутис, у некоторых видов в нем образуются местные окостенения, которые упрочняют кожу. У черепах такие окостенения формируют костный панцирь, состоящий из спинного щита - карапакса, и брюшного - пластрона.
Бедная железами, покрытая толстым ороговевающим эпидермисом кожа рептилий значительно хуже пропускает воду, чем покровы амфибий, особенно экономно испаряют воду через кожу пустынные виды пресмыкающихся (например, серый варан, агамы, круглоголовки и др.). Однако испарение через кожу все-таки идет, например, крокодилы, которым воду экономить не приходится (разумеется, если нет засухи!), теряют таким образом до 75% выделяемой воды.
В отличие от лягушек, кожа рептилий плотно срастается с телом, не образуя лимфатических мешков.
Скелет рептилий (особенно осевой) во многом отличается от скелета амфибий. Позвоночный столб состоит из процельных позвонков (передняя торцевая поверхность вогнутая, а задняя - выпуклая) у высших форм (например, у большинства чешуйчатых) или амфицельных (двояковогнутых) у низших (например, у гаттерий). Более сложная композиция позвонков у черепах: передние позвонки - опистоцельные, средние - амфицельные, а задние - процельные.
Дифференцировка на отделы становится более заметной, поскольку количество позвонков в каждом из них увеличивается. Позвоночный столб подразделяется на четыре отдела: шейный, пояснично-грудной, крестцовый и хвостовой. Шейный отдел всегда насчитывает более одного позвонка (например, у ящериц их восемь), что делает шею (и, соответственно, голову!) значительно более подвижной по сравнению с амфибиями. Это обстоятельство чрезвычайно важно для животного, поскольку с выраженной шеей оно может не только наклонять или поднимать голову, что для позвоночника лягушки является вершиной подвижности, но и поворачивать ее во все стороны, обозревая при этом окрестности.
Не меньшее функциональное значение имеет обособление первых двух шейных позвонков в атлантоосевой комплекс. Об этих позвонках подробно рассказано в разделе, посвященном организации скелета туловища человека (см. т. 1), поэтому здесь мы лишь напомним, что первый шейный позвонок - атлант - посредством верхних суставных поверхностей (у рептилий и птиц такая поверхность одна) подвижно сочленяется суставами с костными выпуклостями - мыщелками затылочной кости. Другой особенностью этого позвонка является то, что у него отсутствует тело, которое переместилось к нижележащему второму шейному позвонку - осевому (эпистрофей), образовав его зуб (рис. 313). Передняя дуга атланта образует с этим зубом подвижный цилиндрический сустав, благодаря чему сам атлант вместе с черепом поворачивается вокруг зуба. Таким образом, у амниот имеется возможность поворачивать голову вокруг продольной оси тела (у человека в связи с прямохождением продольная ось тела совпадает с его вертикальной осью). Обращаем внимание, что у рептилий череп соединяется с атлантом посредством лишь одного затылочного мыщелка.
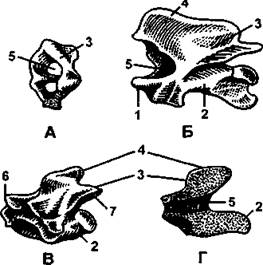
Рис. 313. Позвонки варана:
А - атлант; Б - эпистрофей; В - грудной позвонок; Г - продольный разрез грудного позвонка; 1 - зубовидный отросток эпистрофея; 2 - тело позвонка; 3 - верхняя дуга; 4 - остистый отросток; 5 - канал для спинного мозга; 6 - передний сочленовный отросток; 7 - задний сочленовный отросток (по H. Н. Карташеву и соавт.)
Грудные позвонки у рептилий внешне не отличаются от поясничных, поэтому эти два отдела позвоночного столба объединяют в один - пояснично-грудной. Морфологически этот отдел отличается от других тем, что к каждому позвонку прикрепляется пара ребер. У ящерицы насчитывается 22 пояснично-грудных позвонка, все они несут ребра, но только первые пять пар снизу присоединяются к грудине, образуя настоящую грудную клетку (у ящериц последние три шейных позвонка также несут по паре хорошо выраженных шейных ребер, однако они не доходят до грудины). Остальные ребра не срастаются с грудиной и своими свободными концами заканчиваются в мышцах стенки тела. Значительно меньше ребер у черепах - всего восемь пар, причем все они срастаются с расположенными выше пластинами карапакса. У змей ребра с грудиной не срастаются вообще, что связано с ползающим образом жизни. Напомним, что у земноводных короткие ребра не срастаются с грудиной, поэтому у них грудная клетка не оформлена.
Крестцовый отдел у пресмыкающихся представлен двумя (а не одним, как у земноводных) позвонками, к их длинным поперечным отросткам присоединяются подвздошные кости тазового пояса.
Позвонки хвостового отдела не имеют подвижно сочлененных ребер, постепенно их размеры уменьшаются по направлению к концу тела. Обычно это самые многочисленные позвонки, но их количество у разных видов широко варьирует и во многом зависит от длины хвоста. У гаттерии, некоторых ящериц, а также у многих вымерших рептилий посередине тела хвостового позвонка во фронтальной плоскости (напомним, что она перпендикулярна продольной оси тела животного) имеется хрящевая неокостеневающая прослойка, которая разделяет тело позвонка на переднюю и заднюю половины. В случае опасности местное рефлекторное сокращение мускулатуры хвоста переламывает тело позвонка по линии хрящевой прослойки, при этом часть хвоста, расположенная каудальнеє, отламывается и продолжает беспорядочно изгибаться за счет сокращений мышц, что вводит в заблуждение врага и дает возможность ящерице благополучно улизнуть. В дальнейшем хвост регенерирует, правда, в несколько ущербном виде. Такой способ адаптивного «самокалечения» называется аутотомией. У черепах все позвонки, кроме шейных и хвостовых, неподвижно срастаются с пластинами карапакса.
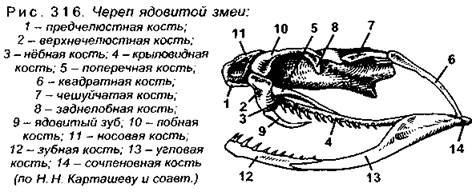
Череп характеризуется почти полным окостенением первичного (хрящевого) черепа, или хондрокраниума, и наличием кнаружи от него большого количества покровных костей, входящих в состав дна, крыши и боковых стенок (рис. 314). Затылочную область мозгового черепа формируют четыре затылочные кости: две боковые, верхняя и основная, которые сообща ограничивают затылочное отверстие. Единственный затылочный мыщелок образуют боковые и основная затылочные кости. Слуховая область состоит из трех пар ушных костей, из которых верхние (верхнеушные) срастаются с верхней затылочной костью, а задние (заднеушные) - с боковыми затылочными, и только передние (переднеушные) кости остаются самостоятельными.
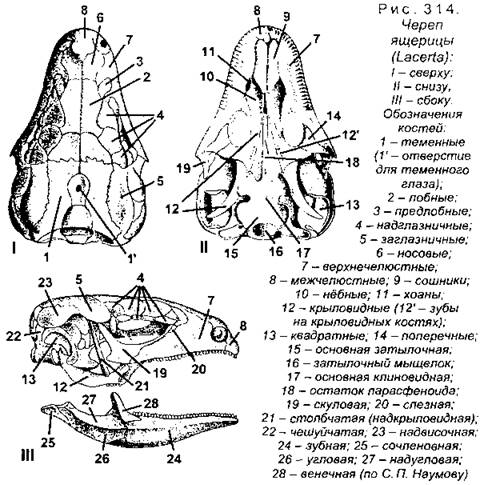
Большую часть основания черепа у рептилий (у всех других амниот тоже) образует покровная основная клиновидная кость, лежащая впереди основной затылочной кости (эти кости срастаются), напомним, что у анамний эту функцию выполняет парасфеноид, который у высших позвоночных отсутствует. Кроме клиновидной, в образовании дна черепа участвуют парные кости: крыловидные, квадратные и нёбные. Крышу черепа образует ряд парных покровных костей: носовые, лобные и теменные (у ящериц они срастаются в непарную кость с отверстием в центре для теменного органа). Большое количество парных покровных костей входит в состав боковых стенок: надвисочные, чешуйчатые, скуловые, предлобные, предглазничные, слезные.
У разных пресмыкающихся мозговой череп образует одну или две височные дуги, например, у крокодила кнаружи от теменных костей с каждой стороны имеется верхняя височная яма. Она отграничена по наружному краю заднелобной (заглазничной) и чешуйчатой костями, которые вместе составляют верхнюю височную дугу. Кроме того, по бокам черепа сзади глазниц располагаются боковые височные ямы, которые снаружи отграничены нижними височными дугами, образованными скуловой и квадратно-скуловой костями (рис. 315). Череп с двумя парами височных дуг и двумя парами височных ям называется диапсидным, или двудужным.
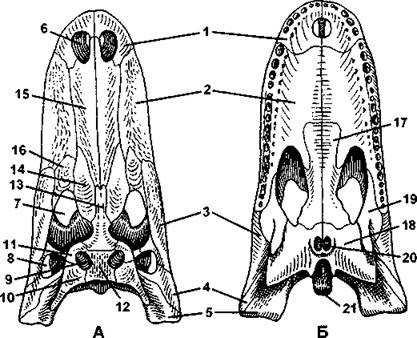
Рис. 315. Череп крокодила (миссисипский аллигатор):
А - сверху: Б - снизу: 1 - межчелюстная кость; 2 - верхнечелюстная кость; 3 - скуловая кость; 4 - квадратно-скуловая кость; 5 - квадратная кость; 6 - наружная ноздря; 7 - глазница; 8 - боковая височная яма; 9 - верхняя височная яма; 10- чешуйчатая кость; 11 - заднелобная (заглазничная) кость; 12 - теменная кость; 13 - лобная кость; 14 - предлобная кость; 15 - носовая кость; 16 - слезная кость; 17 - нёбная кость; 18 - крыловидная кость; 19 - поперечная кость; 20 - хоаны (внутренние отверстия ноздрей); 21 - затылочный мыщелок (по H. Н. Карташеву и соавт.)
У некоторых ящериц сохраняются полные верхние височные дуги, отделяющие верхние височные ямы, тогда как в нижней дуге сохраняется только скуловая кость, а квадратно-скуловая кость отсутствует, из-за чего боковые височные ямы остаются снаружи незамкнутыми (т.е. открытыми). У других ящериц частично редуцируются не только нижние, но и верхние дуги, а у змей обе височные дуги вообще отсутствуют (заднелобная и чешуйчатая кости не соединяются) и обе височные ямы остаются открытыми снаружи (рис. 316). Височные ямы весьма полезны, т.к. позволяют в значительной мере снизить массу черепа.
Наконец у черепах височные ямы отсутствуют (рис. 317), соответственно, их череп называется анапсидным, или бездумным.
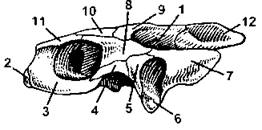
Рис. 317. Череп болотной черепахи:
1 - ложная височная яма; 2 - предчелюстная кость; 3 - верхнечелюстная кость; 4 - скуловая кость; 5 - квадратно-скуловая кость; 6 - квадратная кость; 7 - чешуйчатая кость; 8 - заднелобная кость; 9 - теменная кость; 10 - лобная кость; 11 - предлобная кость; 12- верхняя затылочная кость (по H. Н. Карташеву и соавт.)
У крокодилов и некоторых черепах имеется вторичное костное нёбо (см. рис. 315), которое образуют нёбные отростки межчелюстных, верхнечелюстных, нёбных и крыловидных костей, сросшиеся по срединной линии. Твердое нёбо разделяет носовую и ротовую полости.
Висцеральный череп характеризуется окостенением нёбно-квадратного хряща, который становится упоминаемой выше квадратной костью. Верхний конец этой кости подвижно сочленен с мозговым черепом, а нижний - с нижней челюстью. Впереди от квадратной кости (напоминаем, что она по происхождению относится к первичному хондральному черепу) располагаются покровные кости: крыловидная и впереди нее нёбная, которые соединяются с сошником и верхнечелюстными костями. От крыловидной кости вверх отходит парная верхнекрыловидная кость, соединяющая крыловидную кость с теменной (она имеется у ящериц и гаттерий), и поперечные кости, которые впереди соединяются с верхнечелюстными костями.
Функционирующая вторичная верхняя челюсть образована предчелюстными (межчелюстными) и верхнечелюстными костями. Нижняя челюсть образована сочленовной костью (относится к первичному черепу) и рядом покровных костей: зубной, угловой, надугловой, венечной и некоторых других (в зависимости от вида). Зубы располагаются на предчелюстной, челюстной, зубной, а также могут находится на крыловидных костях. Все зубы имеют одинаковое строение, поэтому жевать рептилии не могут и вынуждены заглатывать пищу целиком. У черепах зубы отсутствуют, вместо них функционируют острые роговые чехлы, покрывающие челюстные кости. У змей кости черепа имеют многочисленные подвижные соединения (рис. 318), благодаря чему они могут проглатывать добычу, которая значительно превосходит по толщине саму змею (например, птичьи яйца).
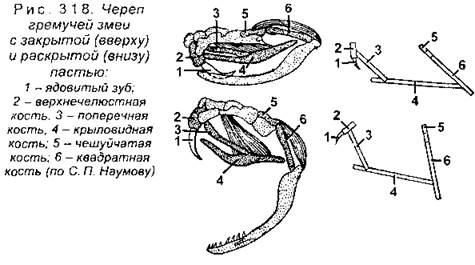
Подъязычная дуга, как и у амфибий, преобразуется в подъязычный аппарат, а гиомандибуляре (подвесок) функционирует как слуховая косточка - стремечко.
Добавочный скелет, в основном, сходен со скелетом амфибий, но отличается некоторыми особенностями. В плечевом поясе имеются ключица (отсутствует у крокодилов) и надгрудинник, которые повышают прочность соединения правой и левой сторон плечевого пояса (рис. 319). Соединение ребер с грудиной и формирование грудной клетки обеспечивает непосредственное соединение плечевого пояса с осевым скелетом (напомним, что у амфибий плечевой пояс располагается в мышцах тела и с осевым скелетом не связан). У змей пояс верхних конечностей редуцируется, а у черепах ключицы и надгрудинник срастаются с пластроном, образуя некоторые его пластинки (передние парные и расположенную между ними непарную).
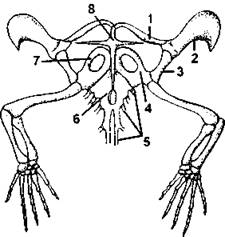
Рис. 319. Плечевой пояс и передние конечности ящерицы (Lacerta):
1 - ключица; 2 - надлопаточный хрящ; 3 - лопатка; 4 - коракоид; 5 - ребра; 6 - грудина; 7 - переднекоракоидный хрящ; 8 - надгрудинник (по С. П. Наумову)
Тазовый пояс, образований подвздошной, седалищной и лобковой костями, не имеет существенных особенностей (рис. 320). Свободные передняя и задняя конечности имеют типичное строение, в области коленного сустава (сочленение бедра с голенью) расположена мелкая сеса- мовидная косточка - коленная чашечка. Проксимальный ряд костей предплюсны срастается с берцовыми костями (или малоподвижно соединяется с ними), а кости дистального ряда также прочно соединяются с костями плюсны, поэтому вместо голеностопного сустава выражен межпредплюсневый сустав, соединяющий проксимальный и дистальный ряды костей предплюсны.
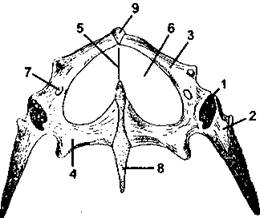
Рис. 320. Тазовый пояс живородящей ящерицы (вид снизу):
1 - суставная впадина для головки бедра; 2 - подвздошная кость; 3 - лобковая кость; 4 - седалищная кость; 5 - связка; 6 - «окно»; 7 - запирательное отверстие; 8 - задний хрящевой отросток; 9 - передний хрящевой отросток (по С. П. Наумову)
У гаттерий и крокодилов вдоль брюха под кожей располагается ряд тонких покровных костей, которые называют брюшными ребрами, однако к настоящим хондральным ребрам они никакого отношения не имеют. Полагают, что они представляют собой остатки брюшного панциря стегоцефалов.
Возможно, костные пластинки пластрона черепах также гомологичны брюшным ребрам.
Мышечная система сохраняет сегментированный характер только в области позвоночного столба, остальная часть скелетной мускулатуры представлена дифференцированными мышцами, расположенными вокруг суставов, которые обеспечивают частные движения определенных костей. Поэтому движения рептилий более сложные и многообразные, чем у амфибий.
Мышцы пояса задней конечности главным образом прикрепляются не к костям таза, а к хвостовым позвонкам. Скорость передвижения пресмыкающихся по твердому субстрату значительно более высокая, чем у амфибий. Увеличение числа шейных позвонков сопровождается дифференцировкой хорошо развитой мускулатуры шеи, из-за чего голова становится подвижной. Поскольку у рептилий имеется полноценная грудная клетка, у них развивается межреберная мускулатура, которая обеспечивает изменения объема грудной клетки, необходимые для реберного типа дыхания.
Пищеварительная система рептилий более дифференцировании, чем у земноводных. Ротовая полость четко отделена от глотки, причем у крокодилов и черепах ротовая полость отделена от носовой посредством костного нёба (см. рис. 315). Железы ротовой полости развиты лучше, чем у амфибий.
На дне ротовой полости имеется подвижный язык, снабженный собственной мускулатурой, причем форма языка у разных видов неодинакова. В частности, у некоторых ящериц (например, агам) он плоский и суживается кпереди, у змей и многих ящериц (например, варанов) язык длинный, тонкий и раздваивается на конце, у хамелеонов очень длинный язык имеет на конце расширение и т.д. Обычно язык участвует в процессе добывания пищи, например, хамелеон с помощью языка захватывает пищу, однако нередко его функция не ограничивается этим, так, змеи и многие ящерицы с помощью языка определяют запахи (в этом участвует якобсонов орган).
У большинства рептилий имеются конические зубы, с помощью которых животное удерживает пищу, однако жевать рептилии не могут, поэтому для того, чтобы оторвать кусок от крупной добычи, приходится, ухватившись за нее зубами, поворачиваться вокруг своей оси (так поступают, например, крокодилы). Зубы располагаются на межчелюстных, верхнечелюстных, нижнечелюстных (зубных) и крыловидных костях, в отличие от амфибий, сошник не несет зубов (исключение составляет гаттерия). Обычно зубы попросту прирастают к кости, и только у крокодилов они находятся в альвеолах, что делает их более устойчивыми.
Пищевод более длинный, чем у земноводных, что объясняется значительно большей длиной шеи рептилий (рис. 321). Напомним, что по пищеводу пища только продвигается из глотки в желудок, который лежит в передней части брюшной полости. Стенки желудка снабжены хорошо развитой мускулатурой, что обеспечивает его перистальтику. Из желудка химус попадает в тонкую кишку, начинающуюся двенадцатиперстной кишкой, в которую впадают протоки печени и поджелудочной железы, расположенной в первой петле тонкой кишки.
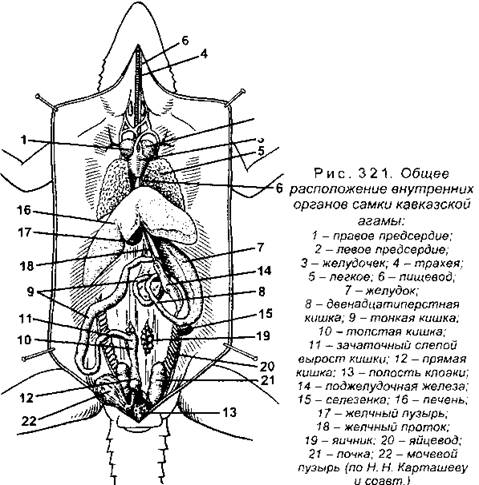
Сделав несколько петель, тонкая кишка переходит в толстую, причем в области перехода у рептилий имеется небольшой вырост - зачаточная слепая кишка, которая у сухопутных растительноядных видов черепах довольно хорошо развита. Напомним, что слепая кишка отсутствует у анамний. Задний отдел толстой кишки образован прямой кишкой, которая открывается в клоаку.
По характеру питания рептилии очень разнообразны, среди них имеются большое количество хищников, например все змеи, вараны, крокодилы и др. Часть видов питается растительной пищей, например агамы, многие черепахи. У рептилий относительно медленный обмен веществ, поэтому они способны длительное время голодать (несколько месяцев и даже более года). Общая длина пищеварительного тракта зависит от характера пищи: у плотоядных он короче, а у растительноядных - длиннее.
Дыхательная система совершеннее, чем у амфибий, что проявляется в более четкой дифференцировке воздухоносных путей и значительном увеличениии функциональной поверхности газообмена в легких. Воздухоносные пути четко подразделяются на верхние - носовую полость (напомним, что у большинства видов она объединена с ротовой полостью, но у крокодилов и черепах эти полости разделены костным нёбом) и нижние - гортань, трахею и бронхи.
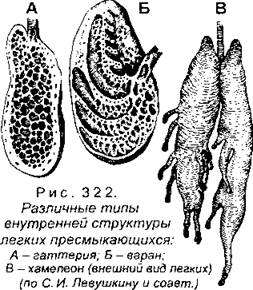
Стенки гортани поддерживают три хряща - парные черпаловидные и непарный перстневидный. Длинная трахея поддерживается в открытом состоянии благодаря кольцевидным хрящам в ее стенках. Легкие имеют разнообразную форму (рис. 322) и более сложное, чем у амфибий, строение. В отличие от последних, на внутренней поверхности легких рептилий образуются не мелкие выросты, а сложная сеть перегородок, которые формируют множество небольших внутренних ячеек, имеющих в совокупности гораздо большую поверхность, чем у мешковидного легкого амфибий. Стенки ячеек пронизаны большим количеством кровеносных сосудов, приносящих венозную кровь для газообмена с легочным воздухом. У многих ящериц (особенно у хамелеонов) задняя часть легких вытянута в виде пальцевидных выростов (они называются легочными мешками), там отсутствуют ячейки, поэтому газообмен не идет. Наиболее простое строение имеют легкие гаттерий, напоминающие легкие амфибий (см. рис. 322), а самое сложное - черепах и крокодилов.
Поступление и удаление воздуха у рептилий происходит не за счет заглатывания, а посредством ритмичного сокращения межреберных мышц, изменяющих объем грудной клетки. При вдохе увеличивается объем грудной клетки, легкие пассивно расширяются, что приводит к уменьшению давления внутри них, поэтому атмосферный воздух через ноздри засасывается в верхние пути, из них через гортанную щель поступает в гортань, затем в трахею, которая делится на два бронха, несущих воздух непосредственно в легкие.
При выдохе грудная клетка сжимается и выдавливает из легких воздух, который удаляется тем же путем, но в обратном направлении. Такой тип дыхания называется реберным (напомним, что у амфибий отсутствует оформленная грудная клетка, поэтому реберное дыхание у них невозможно).
Кожное дыхание у рептилий отсутствует, поскольку покровы тела имеют толстый роговой слой, который очень сильно ограничивает свободную диффузию газов.
Поскольку эмбриональное развитие пресмыкающихся полностью происходит в яйце и не связано с естественными водоемами, жаберное дыхание у них отсутствует, а газообмен в яйце осуществляется за счет алантоиса и желточного мешка.

Рис. 323. Сердце ящерицы:
1 - общий ствол сонных артерий; 2 - внутренняя сонная артерия; 3 - наружная сонная артерия; 4 - легочная артерия; 5 - левая дуга аорты; 6 - правая дуга аорты; 7 - подключичная артерия; 8 - легочная вена; 9 - полая вена (нижняя) и две яремные вены (верхние); 10 - спинная аорта; 11 - желудочно-кишечная артерия (к внутренностям) (по С. П. Наумову)
Кровеносная система рептилий более дифференцирована, что позволяет эффективнее разделять артериальную и венозную кровь. Сердце состоит из трех камер, но предсердия разделены полной перегородкой, при этом предсердно- желудочковое отверстие не общее, как у амфибий, а индивидуальное для каждого предсердия. Венозный синус входит в состав правого желудочка, артериальный конус редуцирован (сохраняется в рудиментарном виде у гаттерий, а также некоторых видов ящериц и черепах), поэтому артерии - правая и левая дуги, а также легочная артерия от желудочка отходят самостоятельно (рис. 323).
Строение желудочка значительно сложнее, чем у амфибий. У всех рептилий желудочек имеет внутренние перегородки, которые препятствуют смешиванию в его полости артериальной и венозной крови. Неполная горизонтальная перегородка разделяет полость желудочка на две камеры - вентролатеральную (легочную), направленную вниз и вправо, и дорзолатеральную, которая занимает верхнюю и левую от перегородки часть желудочка. Дорзолатеральная камера делится крупным мышечным гребнем, называемым вертикальной перегородкой, на две части - левую (артериальную) и правую (венозную), которая сообщается с вентролатеральной камерой.
В систоле предсердий кровь из них поступает в желудочек через отдельные отверстия, при этом артериальная кровь из левого предсердия выталкивается в левую часть дорзолатеральной камеры желудочка, а венозная кровь из правого предсердия - в правую часть той же камеры. Из правой части кровь затем перетекает в вентролатеральную камеру, с которой, как мы уже говорили, она сообщается. Смешиванию крови в этой камере препятствуют предсердно-желудочковые клапаны, которые в это время прилегают к вертикальной перегородке желудочка, поэтому венозная и артериальная части дорзолатеральной камеры оказываются разделенными.
Артериальная система большого круга кровообращения. Когда начинается систола желудочка и давление в его полости возрастает, венозная кровь из легочной (вентролатеральной) камеры, объединенной с венозной частью дорзолатеральной камеры, поступает в легочную артерию, с которой начинается малый круг кровообращения (рис. 324). От артериальной части дорзолатеральной камеры с левой стороны желудочка отходит правая дуга аорты, несущая артериальную кровь, а от середины желудочка - левая дуга аорты, в которой кровь, казалось бы, должна была быть венозной. Однако давление крови в момент систолы отодвигает перегородку, разделяющую артериальную и венозную камеры желудочка, поэтому в венозную камеру также поступает чистая артериальная кровь, хотя иногда в левую дугу аорты может поступать и смешанная кровь.
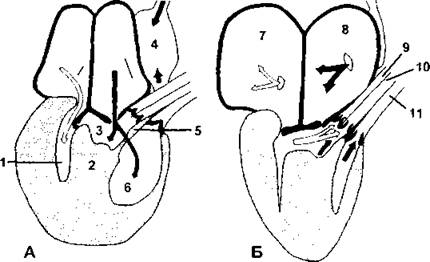
Рис. 324. Схема циркуляции крови в сердце варановой ящерицы.
Представленная картина характерна не для всех ныне живущих пресмыкающихся, весьма многообразных в отношении способов циркуляции: А - сердце в момент сокращения (систолы) предсердий и расслабления (диастолы) желудочка; Б —в момент диастолы предсердий и систолы желудочка. Белыми стрелками показаны потоки артериальной крови, а черными - потоки венозной. В данном случае желудочек частично подразделен вертикальной перегородкой (2) на артериальную полость (1) и второй объем, который также подразделен другой неполной перегородкой, часто именуемой мышечным гребнем (5), на венозную полость (3) и легочную полость (6). На А черная стрелка обходит свободный край мышечного гребня: 4 - венозная пазуха; 7 - левое предсердие; 8 - правое предсердие; 9 - правая системная дуга; 10 - левая системная дуга; 11 - легочная артерия (по Веббу и соавт.)
Поскольку левая дуга аорты отходит от желудочка правее правой дуги, эти сосуды образуют характерный для рептилий перекрест. В дальнейшем от правой дуги отходят сонные и подключичные артерии, несущие артериальную кровь к голове и обеим передним конечностям. Обе дуги аорты (правая и левая) огибают сердце с боков и ниже его сливаются в непарную спинную аорту, от которой отходят ветви ко всем внутренним органам и стенкам тела. В задней части тела аорта отдает две крупные подвздошные артерии (они кровоснабжают задние конечности) и продолжается в хвостовую артерию.
Организация венозной системы не имеет принципиальных отличий от амфибий, за исключением отсутствия кожных вен, несущих артериальную кровь (напомним, что у рептилий отсутствует кожный газообмен).
Малый круг кровообращения начинается с легочной артерии, которая вскоре после отхождения от желудочка делится на две ветви, направляющиеся в правое и левое легкое, где распадаются на капилляры. Артериальная кровь оттекает от легких по правой и левой легочным венам, которые сливаются в единую вену, впадающую в правое предсердие.
Более сложно устроено сердце у крокодилов, поскольку у них имеется полная перегородка, разделяющая полость желудочка на две изолированные части. Поэтому артериальная кровь попадает в правую дугу, а в левую течет венозная кровь, однако у них в основании артериальных стволов имеется проход (паниццево отверстие), через который артериальная кровь все же проникает из правой дуги в левую. Итак, у крокодилов артериальная кровь почти отделена от венозной.
По сравнению с амфибиями рептилии имеют большую частоту сердцебиений (например, у прыткой ящерицы примерно 65 сокращений в минуту), более высокий сердечный индекс (до 2,1), у них выше давление крови и быстрее кровоток. Кроме того, в крови рептилий содержится в два раза больше эритроцитов и значительно больше кислородная емкость крови (см. табл. 5). Все это делает интенсивность обмена веществ примерно в 5 - 10 раз более высокой, чем у земноводных.
Лимфатическая система развита хорошо. Лимфатические сердца немногочисленны, самые крупные из них расположены в области таза. Лимфатические сосуды впадают в различные вены, но, главным образом, в яремные вены.
Выделительная система у всех амниот, в том числе и пресмыкающихся, представлена гораздо более совершенной тазовой почкой, или метанефросом. Характерная для взрослых анамний туловищная почка (мезонефрос) у амниот функционирует только определенный период эмбрионального развития, а потом заменяется метанефросом, который функционирует у взрослой особи в течение всей жизни. Таким образом, у эмбрионов амниот последовательно сменяют друг друга три поколения почек: пронефрос, мезонефрос и метанефрос, образующиеся из общего тяжа нефрогенной ткани, причем каждая из них (начиная с мезонефроса) образуется позади предыдущей почки.
Морфофункциональной единицей метанефроса является нефрон, состоящий из почечного тельца (капсула Шумлянского - Боумена с сосудистым клубочком), в котором фильтруется первичная моча, и системы канальцев, где происходит обратная реабсорбция и формирование вторичной мочи. В отличие от нефронов мезонефроса, здесь всегда отсутствует воронка, а канальцы имеют значительно большую длину, поэтому обратное всасывание из первичной мочи веществ, необходимых организму, которые проходят через фильтрационный барьер и в результате оказываются в первичной моче, идет гораздо более эффективно.
Различают проксимальный каналец, отходящий от капсулы почечного тельца, и дистальный, который открывается в собирательную трубочку. При этом в проксимальный каналец поступает относительно большое количество первичной мочи, а из дистального канальца в собирательную трубочку течет вторичная моча, объем которой значительно меньше. В проксимальном канальце происходит активное (с затратами энергии) всасывание из первичной мочи электролитов, глюкозы и других осмотически активных веществ, в результате чего тканевая жидкость вокруг канальца становится гипертоничной и в соответствии с правилами осмотического транспорта всасывает воду из мочи, находящейся в канальцах. В результате объем мочи уменьшается и она из первичной становится вторичной.
Все реабсорбируемые вещества поступают в кровеносные капилляры, которые густо оплетают канальцы нефрона, причем имеет место противоточный механизм, т.е. направление движения мочи в канальце противоположно току крови в сосудах, что повышает эффективность процесса. Следует отметить, что реабсорбции подвергаются только вещества, необходимые организму, тогда как продукты обмена остаются в полости канальца, постепенно концентрируясь.
Нефроны рептилий организованы проще, чем у других амниот, проксимальные канальцы у них непосредственно переходят в дистальные. Количество нефронов в почках рептилий гораздо большее, чем у амфибий (около 5000).
Конечным продуктом азотистого обмена у пресмыкающихся является нерастворимая мочевая кислота, для выведения которой не требуется большого количества воды, что позволяет рептилиям экономить воду. Соли мочевой кислоты секретируются в просвет канальца и в виде суспензии поступают в собирательные трубочки, а из них в мочевыводящие пути. Чтобы кристаллы кислоты не осаждались на стенках, клетки внутренней выстилки мочевыводящих путей выделяют слизь, содержащую муцин, что предохраняет стенки органа от повреждений.
Мочевая кислота как конечный продукт обмена выводится у большинства видов рептилий, имеющих, таким образом, урикотелический тип обмена. Однако у гаттерии выводится не только мочевая кислота, но и мочевина (уреотелический тип), а у водных черепах - мочевина и аммиак (аммониотелический тип обмена).
Из почек моча поступает в мочеточники, которые в ходе эмбрионального развития отшнуровываются от задней части вольфовых протоков. Мочеточники открываются в клоаку со спинной стороны, с брюшной стороны в клоаку открывается мочевой пузырь (отсутствует у крокодилов и змей).
Половая система. Мужская половая система представлена семенниками, половыми протоками, у самцов большинства рептилий имеются совокупительные органы (рис. 325). Парные семенники подвешены на брыжейке в полости тела по бокам от позвоночного столба. У самцов редуцируется не весь пронефрос - сохраняется его передняя часть, которая затем преобразуется в придаток семенника. От семенника отходят многочисленные семявыносящие канальцы, продолжающиеся в канальцы придатка, которые, в свою очередь, открываются в семяпровод - вольфов проток. Мюллеров проток у самцов не сохраняется. Семяпроводы открываются в мочеточник на своей стороне тела незадолго до его впадения в клоаку. Выросты стенки клоаки образуют совокупительный орган, который у ящериц и змей является парным, у черепах и крокодилов непарным, а у гаттерий отсутствует вообще. При половом возбуждении совокупительные органы выпячиваются наружу, один из них во время полового акта вводится в клоаку самки, куда затем вводится сперма.
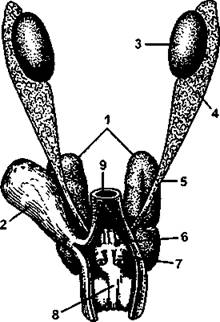
Рис. 325. Мочеполовой аппарат самца кавказской агамы:
1 - почка; 2 - мочевой пузырь; 3 - семенник; 4 - придаток семенника; 5 - семяпровод; 6 - мочеполовое отверстие; 7 - совокупительный мешок; 8 - полость клоаки; 9 - прямая кишка (по H. Н. Карташеву и соавт.)
У черепах и крокодилов вдоль брюшной стенки клоаки проходит пара гребней - пещеристых тел, между которыми имеется канавка, заканчивающаяся головкой. Такая структура очень напоминает половой член млекопитающих. При половом возбуждении головка и пещеристые тела наполняются кровью и значительно увеличиваются. Головка высовывается из клоаки и вводится в клоаку самки, а раздувшиеся пещеристые тела смыкаются свободными краями, превращая канавку между ними в замкнутую трубку, по которой в клоаку самки перетекает сперма. Интересно, что у самок в зародышевом состоянии эти структуры также закладываются, но в дальнейшем не развиваются, а у взрослой самки сохраняются в виде нефункционального клитора.
Женская половая система образована парными яичниками и половыми путями (рис. 326). Яичники лежат в полости тела, будучи подвешенными на брыжейке к стенке тела. Созревшие яйцеклетки, содержащие большое количество желтка, попадают в полость тела и оттуда проникают в воронку одного из яйцеводов, которые одним концом (воронкой) открываются в полость тела, а другим - в клоаку.
В качестве яйцевода у рептилий функционируют мюллеровы протоки (у самок вольфовы протоки не сохраняются), которые иногда подразделяют на отделы. Первый отдел, который начинается воронкой, называется собственно яйцеводом, или маточной трубой, второй - более широкий - маткой, более узкий отдел, соединяющий матку с клоакой, называется влагалищем. Обращаем внимание читателя на то, что, несмотря на общие названия отделов половых путей у рептилий (а также птиц) и млекопитающих, функции этих отделов различны, поэтому названия являются во многом условными.
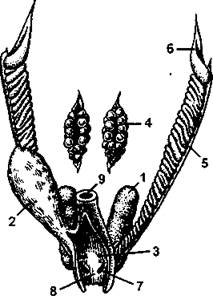
Рис. 326. Мочеполовой аппарат самки кавказской агамы:
1 - почка; 2 - мочевой пузырь; 3 - мочевое отверстие; 4 - яичник; 5 - яйцевод; 6 - воронка яйцевода; 7 - половое отверстие; 8 - полость клоаки; 9 - прямая кишка (по H. Н. Карташеву и соавт.)
Проходя по яйцеводу, яйцеклетка окружается оболочками, выделяемыми клетками эпителия: в верхнем отделе (маточной трубе) — яичным белком, в среднем (матке) - скорлупой, которая у рептилий кожистая, пергаментообразная, иногда пропитана известью. Понятно, что наличие скорлупы требует оплодотворения яйцеклетки до ее образования, поэтому у рептилий оплодотворение внутреннее.
Развитие начинается в яйцеводах и практически полностью завершается в яйце. Напомним, что рептилии являются амниотами, поэтому у них зародыш в яйце окружен оболочками, а скорлупа не дает яйцу высохнуть. Яйцо рептилий содержит много желтка, который скапливается у одного из полюсов (вегетативного). Дробление дискоидальное, при котором борозды дробления не проходят все яйцо, поэтому не охватывают богатую желтком цитоплазму вегетативного полушария, при этом на анимальном полюсе образуется дискообразная однослойная бластодерма, которая затем становится многослойной. Развитие идет без метаморфоза, поэтому из яйца выходит вполне сформированное животное, отличающееся от взрослой особи, главным образом, маленькими размерами. Самка обычно откладывает яйца в подходящих для этого местах. Некоторые виды проявляют заботу о потомстве (например, королевские кобры, которые делают гнезда и охраняют их). Вторичноводные пресмыкающиеся для размножения выходят на сушу и там откладывают яйца (например, морские черепахи) или становятся яйцеживородящими (например, морские змеи).
Нервная система организована сложнее, чем у амфибий, что вызвано адаптацией к постоянно наземному обитанию. В наибольшей степени усложнился головной мозг (рис. 327). Самым крупным отделом является передний мозг, прежде всего, полушария конечного мозга. Основная масса серого вещества полушарий сосредоточена в полосатых телах, расположенных в основании конечного мозга, у рептилий они становятся главными координаторами нервной деятельности.
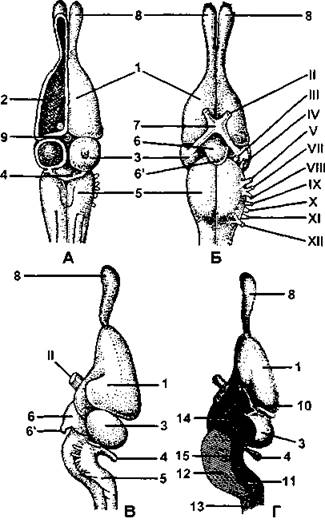
Рис. 327. Головной мозг ящерицы:
А - сверху; В - снизу: В - сбоку; Г - сбоку в разрезе; 1 - полушарие переднего мозга; 2 - полосатое тело; 3 - средний мозг; 4 - мозжечок: 5 - продолговатый мозг; 6 - воронка; 6’ - гипофиз; 7 - хиазма; 8 - обонятельная доля; 9 - эпифиз; 10- промежуточный мозг; 11 - четвертый желудочек; 12- продолговатый мозг; 13- спинной мозг; 14- третий желудочек; 15- сильвиев водопровод; Il - XII - черепные нервы (по Паркеру, с изменениями и дополнениями)
Головной мозг, в котором интегрирующим центром являются полосатые тела переднего (конечного) мозга, относят к зауропсидному типу, кроме рептилий, мозг такого типа имеют также птицы. В полушариях переднего мозга рептилий четко дифференцированы обонятельные доли, кроме того, обособляются теменные доли. Прогрессивной особенностью мозга рептилий является появление у них на поверхности полушарий отдельных участков серого вещества - коры, которые представляют собой зачаток вторичного свода, или неопаллиума. Однако строение коры рептилий очень примитивно, поэтому ее называют древней корой (archicortex).
Крупные полушария конечного мозга прикрывают промежуточный мозг, поэтому сверху его не видно. У рептилий очень хорошо развит теменной орган, строение которого очень напоминает глаз — в нем различаются структуры, сходные с сетчаткой и хрусталиком (рис. 328). Особенно хорошо теменной орган развит у гаттерий, у которых он функционирует в качестве третьего глаза, эффективность такого зрения невелика, но с его помощью животное вполне может отличить свет от тьмы. Гипофиз пресмыкающихся состоит из трех долей (напомним, что передняя и средняя - аденогипофиз, задняя - нейрогипофиз), причем задняя доля развита очень хорошо, что связано с более сложной регуляцией водного обмена у рептилий по сравнению с амфибиями и рыбами.
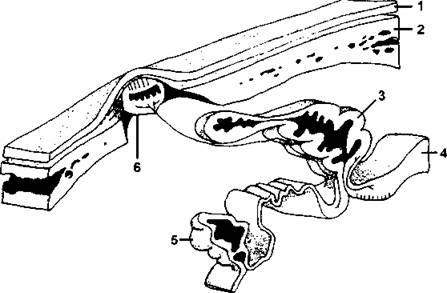
Рис. 328. Теменной глаз и смежные с ним структуры у ящерицы.
Эпифиз (или пинеальный орган) и теменной глаз (париетальный, или парапинеальный, орган) представляют собой дорсальные выросты промежуточного мозга, которые в разных группах позвоночных могут формировать глазоподобные структуры. Парафиз также представляет собой дорсальный вырост, но никогда не напоминает глаз. У некоторых ящериц (и у гаттерий) теменной глаз располагается в теменном отверстии черепа и высокодифференцирован: верхняя стенка этого пузыря уподобляется хрусталику, а нижняя - сетчатке. Он соединяется с крышей промежуточного мозга тонким нервом: 1 - кожа; 2 - крыша черепа; 3 – эпифиз; 4 - средний мозг; 5 – парафиз; 6 - теменной глаз (по Вуртману, с изменениями)
Размеры среднего мозга относительно невелики, поскольку этот отдел не является ведущим. Мозжечок развит значительно лучше, чем у амфибий, поэтому рептилии могут совершать достаточно сложные движения. Продолговатый мозг образует второй изгиб в сагиттальной плоскости, который также имеется у всех остальных амниот (птиц и млекопитающих). Напомним, что первый изгиб расположен в области среднего мозга.
Общая масса головного мозга у рептилий примерно равна массе спинного мозга. Спинной мозг имеет обычное строение, в области отхождения волокон, участвующих в образовании сплетений, образуются утолщения. Как и у амфибий, спинной мозг короче позвоночного столба, поэтому задняя часть спинномозгового канала занята не мозгом, а нервными волокнами и оболочками. Большое значение имеет ретикулярная формация спинного мозга и ствола головного мозга, регулирующая функцию двигательных проводящих путей.
Периферическая нервная система. От головного мозга отходят одиннадцать пар черепных нервов. При этом у рептилий имеется XII пара - подъязычный нерв, который отходит от продолговатого мозга (напомним, что у амфибий соответствующий нерв отходит не от ствола головного мозга, а от спинного мозга, поэтому его нельзя считать черепным нервом). Однако предыдущая - XI пара (добавочный нерв) - соединена с X парой, поэтому общее количество черепных нервов равно одиннадцати.
Спинномозговые нервы образуют характерные для наземных позвоночных сплетения (плечевое и пояснично-крестцовое, или тазовое), волокна которых иннервируют кожу и мышцы конечностей.
Органы, чувств. Орган зрения характеризуется наличием в ресничной мышце поперечнополосатых мышечных волокон, что делает аккомодацию глаза более эффективной, поскольку эта мышца способна не только перемещать хрусталик назад и вперед, но также изменяет его кривизну (напомним, что у рыб аккомодация ограничивается лишь изменением расстояния от хрусталика до сетчатки). В склере глазного яблока имеется кольцо из тонких костных пластинок, а от задней части стенки в заполненную стекловидным телом полость глазного яблока вдается гребешок - вырост, богатый кровеносными сосудами (рис. 329).
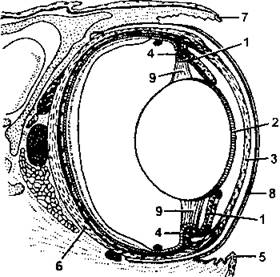
Рис. 329. Глаз пресмыкающегося (продольный разрез глаза змеи):
1 - мышцы радужки; 2 - передняя стенка хрусталика; 3 – роговица; 4 - реснитчатое тело с мышцами; 5 - подглазничная чешуя; 6 – склера; 7 - надглазничная чешуя; 8 - сросшиеся веки; 9 - циннова связка
В отличие от амфибий, глазные яблоки рептилий не могут втягиваться (напомним, что у земноводных глаза втягиваются при глотании пищи и механически проталкивают ее в пищевод), но способны вращаться с помощью глазодвигательных мышц. Особенно эффектно это получается у хамелеонов, глаза которых не только весьма подвижны, но при этом движутся независимо друг от друга. У большинства рептилий имеются веки (верхнее, нижнее и третье в виде мигательной перепонки), однако у гекконов и змей подвижных век нет (у змей они сращены между собой), поэтому гекконы попросту облизывают время от времени роговицу, а змеи периодически удаляют поверхностный слой сросшихся век во время линьки.
Орган обоняния развит очень хорошо, он более сложен, чем у амфибий. Имеются наружные и внутренние ноздри, сообщающие ротовую полость с внешней средой. В носоглоточном ходе обонятельный отдел отделяется от расположенного ниже дыхательного отдела посредством особого выроста - челюстной раковины, передняя часть хода образует некоторое расширение - преддверие (рис. 330).
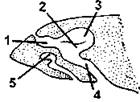
Рис. 330. Схема строения обонятельного мешка ящерицы:
1 - преддверие; 2 - дыхательный отдел; 3 - обонятельный отдел; 4 - носоглоточный ход; 5 - якобсонов орган (по Шимкевичу)
У ящериц и змей хорошо развит якобсонов орган, полость которого сообщается с верхней стенкой полости рта специальным каналом. Чтобы задействовать этот орган, ящерица периодически высовывает язык, который контактирует с летучими веществами воздуха и частицами грунта, при этом часть молекул попадает в слюну, покрывающую язык. После этого животное втягивает язык и прижимает его к каналу якобсонова органа, который анализирует химический состав веществ. Таким образом, пресмыкающиеся способны хорошо различать запахи.
Орган слуха; как и у амфибий, состоит из среднего и внутреннего уха, при этом в перепончатом лабиринте обособляется улитка. У некоторых змей между ноздрями и глазами располагаются парные ямки, с помощью которых животное способно различать инфракрасное излучение живой добычи, даже не видя ее. Эффективность органа очень высока, что позволяет улавливать разницу температур до тысячных долей градуса. Змеи, которые обладают таким термолокатором, называются ямкоголовыми (например, гремучие змеи).
Поведение пресмыкающихся намного сложнее, чем амфибий, что связано с более высокой организацией нервной системы, прежде всего, переднего мозга. При этом, наряду с врожденными рефлексами (инстинктами), большое значение приобретает условно-рефлекторная деятельность, которая формируется в течение жизни, она обеспечивает пластичную адаптацию к резким изменениям условий обитания.