Большинство представителей надкласса рыб относятся к классу костные рыбы. В настоящее время известно около 25 000 современных рыб, однако ихтиофауна больших глубин еще исследована недостаточно полно, поэтому многие виды до сих пор неизвестны людям.
Класс костные рыбы подразделяется на четыре подкласса: хрящекостные (Chondrostei), лучеперые (Actinopterygii), кистеперые (Crossopterygii) и двоякодышащие (Dipnoi).
Скелет костных рыб образован костями, однако у некоторых примитивных видов (подкласс хрящекостных, к которым относятся осетровые рыбы), наряду с костями, присутствуют и хрящевые элементы скелета.
Внешнее строение. Как и у хрящевых рыб, тело костных рыб подразделяется на голову, туловище и хвост. Отсутствие шеи, хорошо обтекаемое гладкое тело без выступов способствует эффективному движению в воде, которая, как известно, значительно плотнее воздуха. Однако образ жизни рыб непременно сказывается на их внешнем виде, в связи с чем выделяют по меньшей мере 12 различных морфологических типов, наиболее распространенные из них это торпедовидный (лучшие пловцы, например тунец, сельдь, треска и др.), змеевидный (угри), лентовидный (относительно плохие пловцы, например рыба-сабля, сельдяной король), стреловидный (способны к резкому ускорению, например, многие хищники - щука, таймень), сплющенный (симметрично-сжатое с боков тело, например у леща, или несимметрично-сжатое, например у камбалы, все они плохие пловцы) (рис. 257).

Рис. 257. Типы формы тела рыб.
А - стреловидный (сарган); Б - торпедовидный (скумбрия); В - сплющенный с боков, лещевидный (лещ обыкновенный); Г - тип рыбы луны (луна-рыба); Д - тип камбалы (речная камбала); Е - змеевидный (угорь); Ж - лентовидный (сельдяной король); З - шаровидный (кузовок); И - плоский (скат) (A-З- костные, И - хрящевая рыбы) (по Б. С. Матвееву и совет.)
На голове располагаются глаза (обычно по бокам головы, но у некоторых видов происходит вторичное смещение наверх или на одну из сторон тела), впереди них находятся обонятельные отверстия, но не на нижней стороне головы (как у акул и скатов), а на верхней. Брызгальце сохраняется только у осетровых. Жаберные дуги находятся в жаберной полости и прикрыты костными жаберными крышками.
Туловищный и хвостовой отделы снабжены кожистыми выростами с костными лучами внутри, которые, как и у хрящевых рыб, могут быть парными или непарными. Парные плавники соответствуют конечностям наземных животным (естественно, иного строения), они выполняют функцию органов движения и главное - рулей, поддерживающих тело в нужном положении и направляющих его во время движения. Строение и расположение плавников зависит от образа жизни рыбы. Например, хорошо развитые грудные плавники позволяют некоторым рыбам уверенно ползать по суше, а летучим рыбам пролетать по воздуху значительные расстояния (до 800 м). Брюшные плавники могут смещаться далеко вперед (например, у тресковых) или видоизменяться в присоску (как это происходит, например, у бычков).
К непарным плавникам относятся спинной, анальный и хвостовой. У некоторых рыб имеются дополнительные плавники, например, у трески на спине располагаются три плавника, у других также могут быть добавочные анальные плавники. У скумбриевых за спинным и анальным плавниками находятся многочисленные добавочные плавнички. У некоторых видов (например, лососевых, хариусовых, корюшковых и др.) за спинным плавником находится еще один, заполненный жировой тканью, но лишенный костной опоры. Эти плавники также могут изменяться, например, спинной плавник марлина имеет очень большие размеры (1,5 м в высоту) и превращается в парус, возвышающийся над водой. Тот же плавник у рыбы-прилипало видоизменяется в присоску. У некоторых видов (например, скорпен) плавники имеют ядовитые шипы. Основной функцией непарных плавников является стабилизация тела в пространстве.
Хвостовой плавник является главным движителем рыбы. Форма хвостового плавника может быть различной (рис. 258), обычно она соответствует скорости плавания. У разных видов в ходе приспособления к определенному образу жизни некоторые плавники могут редуцироваться. Например, у угрей отсутствуют брюшные плавники, а у мурены - грудные.

Рис. 258. Различные типы хвоста: А - симметричный; Б - несимметричный (гетероцеркальный); В - ложносимметричный (по Никольскому)
Особенности строения тела определяют способ плавания рыбы. Большинство видов передвигаются за счет боковых колебательных движений задней части тела и хвоста (рис. 259). Другие перемещаются посредством колебательных изгибов всего тела во фронтальной плоскости. Последний способ более медлительный и характерен для придонных рыб с удлиненным телом (например, угрей).

Рис. 259. Схема локомоции рыб. Стрелками обозначены направления сил, в сумме формирующих поступательное движение (по С. И. Левушкину и соавт.)
Анатомия. Покровы тела, так же как и у хрящевых рыб, представлены кожей с чешуей. Кожа состоит из многослойного эпидермиса и нижележащей плотной дермы. В эпидермисе имеются многочисленные одноклеточные железы, выделяющие слизь (рис. 260). Наружный слой эпидермиса оро- говевает, но не отмирает и сохраняет связь с живыми клетками. У многих видов (например, у сиговых, карповых и др.) перед нерестом степень ороговения покровов увеличивается, а затем восстанавливается (рис. 261).
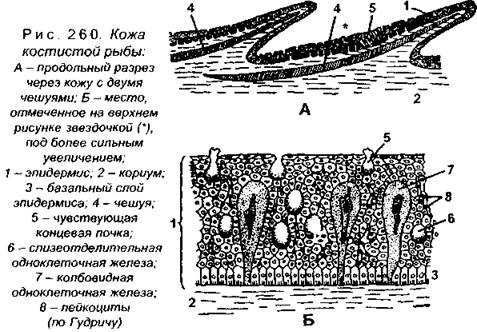

Слизь выделяют эпидермальные клетки трех типов: бокаловидные, зернистые и колбовидные. Установлено, что рыбы с хорошо развитой чешуей (например, окуневые) выделяют меньше слизи, а с редуцированной чешуей или с голой кожей (например, линь, некоторые сомы и др.) - очень много. Слизь выполняет многочисленные функции. Так, в частности, она обладает бактерицидными свойствами и защищает рыбу от патогенных микроорганизмов, участвует в выделении продуктов обмена и водно-солевом обмене, ускоряет свертывание крови и уменьшает трение рыбы о воду. Слизистые клетки эпидермиса синтезируют видоспецифические вещества, позволяющие рыбам узнавать друг друга на расстоянии, причем эти выделения различны не только у отдельных видов, но и у особей одного вида, но разного пола или возраста. При нападении врага из поврежденной кожи (например, у карповых) выделяется ихтиоптерин, который еще называют «веществом страха». У некоторых видов слизь токсична, иногда имеются довольно крупные ядовитые железы, которые так же являются производными эпидермиса, как и светящиеся органы многих глубоководных рыб.
В нижних слоях эпидермиса находятся пигментные звездчатые клетки - хроматофоры. Пигмент может иметь различную окраску, благодаря этому многие виды (например, тропические рыбы, обитающие в коралловых рифах) выглядят очень пестро. Окраска может меняться, что зависит от состояния пигментных клеток - если пигмент собирается в центральной части клетки, большая ее часть становится неокрашенной и цвет тела становится бледнее, если пигмент равномерно растекается по всей цитоплазме, окраска становится ярче (рис. 262). Некоторые виды способны изменять окраску тела довольно быстро, например, лежащая на дне камбала приобретает цвет донного субстрата (рис. 263). Окраска меняется в период размножения (например, у лососевых), при изменении температуры, состава воды, из-за эмоционального состояния рыбы (например, страх) и т.п.

Рис. 262. Меланофор карася: справа - фаза контракции, слева - фаза экспансии: 1 - пигментные зерна; 2 - скелетная фибрилла; 3 - ядра (по Строганову)
Кожа костных рыб покрыта чешуей, однако степень ее развития у разных видов неодинакова (например, у индийского усача она достигает нескольких сантиметров, а у угря почти незаметна), у некоторых рыб (например, у сомов) чешуя может вторично отсутствовать.
Чешуя костных рыб представлена костными чешуйками, которые образуются в кориуме. Наиболее примитивной является ганоидная чешуя, костные пластинки которой покрыты дентинообразным веществом ганоином (ею обладали многие ископаемые формы, из ныне живущих рыб она имеется у панцирной щуки и многоперов, кроме того, у осетровых слившиеся ганоидные чешуи образуют бляшки - жучки), и космоид- ная, покрытая космином (такая чешуя имеется у кистеперых и двоякодышащих рыб). У большинства костных рыб чешуя эласмоидная, т.е. образована только костными пластинками. В зависимости от состояния внешнего края различают циклоидную (гладкий край, например у карповых) и ктеноидную (наружный край покрыт зубчиками, например у окуневых) чешую (рис. 264). Циклоидная чешуя является более примитивной. Иногда у одного вида (например, у лиопсетты из камбаловых) самцы обладают ктеноидной чешуей, а самки - циклоидной. Случается, что чешуи разного типа присутствуют у одной особи.

Рис. 263. Изменение окраски камбалы в зависимости от окраски субстрата (по Б. С. Матвееву и соавт.)

Рис. 264. Форма чешуи рыб: А - ганоидная; Б – циклоидная; В – ктеноидная;1 - годичные кольца (по И. М. Анисимовой, с изменениями и дополнениями)
Чешуи черепицеобразно накладываются друг на друга, образуя ряды, расположение которых и число чешуй в каждом из них характерно для определенного вида, кроме того, этот показатель не меняется с возрастом рыбы. Рост чешуек в течение года идет неравномерно, поэтому на них образуются годичные слои и по ним можно определить возраст исследуемой особи (рис. 265).
Изначально чешуя развилась в качестве защитного приспособления, однако не меньшее ее значение (а может, даже значительно большее) состоит в улучшении гидродинамических свойств рыбы. Известно, что виды с плохо развитой чешуей (или даже без нее) плавают хуже тех, у кого чешуя крупная, несмотря на то, что тело бесчешуйных рыб покрыто более толстым слоем слизи.

Рис. 265. Соотношение между скоростью роста рыбы и ее чешуи (по Никольскому)
В коже располагаются различные рецепторы, поэтому она имеет важное сигнальное значение, кроме того, у костных рыб в дерме образуются покровные кости черепа и кости пояса передних конечностей.
Скелет костных рыб образован костями, лишь у осетровых в течение жизни сохраняется значительное количество хрящевых элементов. Костный скелет эффективнее выполняет свои функции, поскольку он значительно прочнее и менее массивен. У рыб костные элементы могут возникать двумя способами. Во-первых, кость развивается из хряща (первичные, или замещающие, кости), во-вторых, из очагов окостенения в дерме кожи (вторичные, или покровные, кости, их также называют накладными, поскольку они могут накладываться на хрящевые элементы скелета, например, в черепе осетровых). Следовательно, в любом случае кости имеют мезодер- мальное происхождение. Скелет подразделяется на осевой и добавочный (рис. 266). Осевой скелет включает в себя позвоночный столб и череп. В позвоночном столбе выделяют туловищный и хвостовой отделы. Позвоночник образован костными позвонками, которые, как и у хрящевых рыб, являются амфицельны- ми (т.е. их тела двояковогнуты - спереди и сзади). От тела вверх отходят верхние дуги, которые, срастаясь между собой, ограничивают позвоночное отверстие. Отверстия всех позвонков образуют позвоночный канал, через который проходит спинной мозг. Верхние дуги продолжаются в непарный верхний остистый отросток (рис. 267).

Рис. 266. Скелет костистой рыбы (окуня):
1 - кости черепа; 2 - основные элементы спинного плавника; 3, 4- лучи спинного плавника; 5 - последние позвонки, удерживающие хвостовой плавник; 6 - хвостовые позвонки; 7 — основные элементы анального плавника; 8 - туловищные позвонки; 9 - ребра с придатками; 10- кости и лучи брюшного плавника; 11 - кости и лучи грудного плавника; 12- жаберная крышка; 13 - верхняя и нижняя челюсти (по Баклашовой)

Рис. 267. Позвонки судака:
А - туловищный позвонок; Б - хвостовой позвонок; 1 - тело позвонка; 2 - поперечный отросток; 3 - ребро; 4 - верхняя дуга; 5 - верхний остистый отросток; 6 - нижняя дуга; 7 - нижний остистый отросток; 8 - мышечная косточка (по H. Н. Карташеву и соавт.)
Снизу отходят нижние дуги, но лишь у хвостовых позвонков они смыкаются, образуя гемальный канал, где проходят хвостовая артерия и вена. Внизу нижние дуги заканчиваются нижним остистым отростком. В туловищном отделе нижние дуги короткие и не срастаются, к ним прикрепляются ребра. В отличие от хрящевых рыб, ребра которых ограничивают полость тела лишь сверху, у костных ребра длинные и ограничивают полость тела еще и с боков. Последний хвостовой позвонок имеет поверхность для прикрепления опорных структур хвостового плавника, поэтому он часто уплощен, однако нередко этот позвонок удлиняется, загибается вверх, образуя уростилъ. Для каждого вида рыб характерно определенное общее количество позвонков, например, у луны-рыбы их 17, у северной сельди - 57, а у угря - 114. Интересно, что повышение температуры в период эмбрионального развития влечет за собой уменьшение числа позвонков.

Рис. 268. Схема черепа костистой рыбы, жаберная крышка и окопоглазничное кольцо удалены.
Хрящевые кости обозначены мелкими точками:
1 - нижнезатылочная кость; 2 - боковая затылочная; 3 - верхнезатылочная; 4 - ушные; 5 - основная клиновидная; 6 - крылоклиновидная; 7 - глазоклиновидная; 8 - межобонятельная; 9 - боковая обонятельная; 10 - теменная; 11 - лобная; 12 - носовая; 13- парасфеноид; 14 - сошник; 15 — нёбная; 16 - квадратная; 17 - крыловидные; 18 - межчелюстная; 19- верхнечелюстная; 20 - сочленовная; 21 - зубная; 22 - угловая; 23 - гиомандибулярная; 24 - симплектикум; 25 - 29 - 1 - 5-я жаберные дуги; 30 - гиоид (по Шмальгаузену)
Череп состоит из мозгового и висцерального отделов (рис. 268). У костных он гораздо более совершенен, чем у хрящевых рыб, и устроен значительно сложнее. Как мы уже говорили, череп частично или полностью становится костным, причем в его составе имеются как первичные (хондральные) кости, так и вторичные (накладные). В процессе онтогенеза вначале образуется первичный череп из хрящевых зачатков, которые затем окостеневают. Одновременно с этим в кориуме формируются накладные кости. Области окостенений первичного черепа соответствуют отделам мозгового черепа. В затылочном отделе образуются четыре затылочные кости (основная, две боковые и верхняя), ограничивающие затылочное отверстие. Ушные кости формируются в области слуховой капсулы, которая расположена впереди затылочного отдела, обычно с каждой стороны имеется по пять таких костей. Стенки глазниц (точнее, межглазничной перегородки) образуют непарная основная клиновидная кость, парные крылоклиновидные и глазоклиновидные кости (эту группу костей, в которых у большинства рыб сохраняются хрящевые элементы, также называют сфеноидами). Самую переднюю часть мозгового черепа (она также содержит хрящевые элементы) - обонятельный отдел (обонятельные капсулы) - составляют непарная срединная обонятельная кость (межобонятельная) и парная боковая обонятельная кость. Эти кости сообща также называются обонятельными, или этмоидами.
Первичные кости образуют заднюю и боковые части мозгового черепа, а также частично его дно, но не охватывают крышу, где, как и у хрящевых рыб, остается большое отверстие - фонтанель. Для более полной защиты головного мозга и органов чувств дополнительно к первичным костям образуются вторичные (накладные) кости, которые составляют основу крыши и входят в состав боковых стенок и дна. В задней части находятся парные теменные кости, впереди от них - парные лобные кости, а перед ними - также парные носовые кости. Боковые отделы черепа включают в себя цепочку небольших глазных косточек (переднюю, самую большую еще называют слезной), в совокупности образующих окологлазничное кольцо. Дно мозгового черепа сформировано, главным образом, непарным парасфеноидом и другой непарной вторичной костью - сошником, расположенным спереди. Кости мозгового черепа соединяются между собой неподвижно, повышая тем самым прочность всей конструкции.
Висцеральный череп составляют челюстная, подъязычная и жаберная дуги (напоминаем, что эти образования имеются и у хрящевых рыб), а также кости жаберной крышки. Строение составляющих висцерального черепа более сложно, чем у хрящевых рыб. Челюстная дуга включает в себя большое количество хондральных и накладных костей. Верхнечелюстной отдел в переднем отделе образован парными нёбными костями, в среднем - тремя крыловидными (две из них являются накладными, а одна хондральной), в заднем - парными квадратными костями. Все они являются производными нёбно-квадратного хряща, описанного ранее у хрящевых рыб. Однако эти первичные кости в большей степени выполняют функцию дна черепа, а непосредственно верхнюю челюсть образуют накладные парные кости: верхнечелюстные и предчелюстные.
Из меккелева хряща, который у хрящевых рыб служит нижней челюстью, образуется первичная сочленовная кость, однако у костных рыб она лишь образует челюстной сустав с квадратной костью верхней челюсти. Основную же часть нижней челюсти составляют парные вторичные кости: крупная зубная кость, которая покрывает часть сочленовной кости, и мелкая угловая кость (она сзади соединяется с сочленовной костью). Таким образом, первичные челюсти костных рыб (т.е. кости, возникшие из нёбно-квадратного и меккелева хрящей) образуют челюстной сустав, тогда как непосредственно челюсти образованы вторичными костями. Наличие вторичных челюстей позволяет значительно разнообразить способы захвата и удержания пищи, приспосабливаться к определенному типу пищи, именно этим можно объяснить различные формы ротового аппарата и, соответственно, видовое разнообразие (рис. 269). Это выгодно отличает костных рыб от хрящевых, ротовой аппарат которых устроен однотипно, и поэтому они не способны потреблять такую разнообразную пищу, как различные виды костных рыб.

Рис. 269. Различные формы строения ротового аппарата рыб (по С. И. Левушкину и совет.)
Подъязычная и жаберные дуги, в общем, образованы теми же элементами, что и у хрящевых рыб, однако все они костные (причем эти кости первичные). Жаберных дуг также пять, но последняя из них подвергается сильной редукции и развита заметно хуже остальных. Подвижное соединение мозгового и висцерального отделов черепа у костных рыб осуществляется при помощи подъязычной дуги, при этом гиомандибуляре образует сустав со слуховым отделом мозгового черепа. Напомним, что такой тип черепа называется гиостилическим.
Жаберная крышка, присущая только костистым рыбам, прикрывает с боков жаберные дуги. Она состоит из четырех плоских накладных костей: крышка, предкрышка, подкрышка и межкрышка (270). Жаберная крышка прикрепляется к гиомандибуляре.
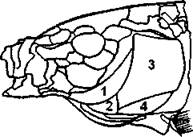
Рис. 270. Кости жаберной крышки: 1 - предкрышка; 2 - межкрышка; 3 - крышка; 4 – подкрышка (по С. И. Левушкину и совет.)
Добавочный скелет образован костями свободных конечностей и их поясов. Пояс передних конечностей - грудных плавников - образован небольшими лопаткой и коракоидом, которые являются первичными костями, и цепочкой вторичных костей, самая крупная из которых - клейтрум, имеющая серповидную форму (эту кость еще называют ключицей), которая соединяется с задним отделом мозгового черепа (рис. 271). У грудных плавников отсутствуют базалии, поэтому костные радиа- лии прикрепляются непосредственно к костям пояса. Скелет самих плавников образован костными лучами кожного происхождения, которые прикрепляются к радиалиям.

Рис. 271. Плечевой пояс и грудной плавник судака:
1 —лопатка; 2 - коракоид: 3 - клейтрум; 4 - надклейтрум: 5 - заднетеменная кость; 6 - заднеключичная кость: 7 - радиапии; 8 - костные кожные лучи (по H. Н. Карташеву и соавт.)
Пояс задних конечностей (тазовый пояс) состоит из двух сросшихся костей, свободно лежащих в толще мышц, к которым прикрепляются костные лучи брюшных плавников (рис. 272). Следовательно, у брюшных плавников отсутствуют не только базалии, но и радиалии.
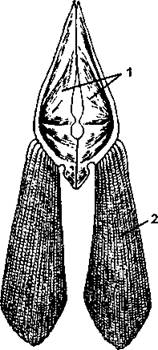
Рис. 272. Тазовый пояс и брюшной плавник судака: 1 - тазовые кости; 2 - кожные костные лучи (по H. Н. Карташеву и совет.)
Непарные конечности поясов не имеют, их опорой являются только костные лучи. Таким образом, добавочный скелет у костных рыб устроен проще, чем у хрящевых.
Кроме описанных костей, опору для соматической мускулатуры образуют многочисленные тонкие межмышечные (их также называют мускульными, или туловищными) косточки, которые находятся в толще мышц. По своему происхождению они являются окостеневшими сухожилиями.
Мышечная система костных рыб развита лучше, чем у хрящевых, что позволяет им совершать значительно более сложные движения. Она представлена мышцами туловища, головы и плавников. Основную массу составляет мускулатура туловища, которая так же, как и у хрящевых рыб, отчетливо сегментирования и состоит из расположенных вдоль тела толстых мышечных массивов, разделенных прослойками соединительной ткани - миосептами на S-образно изогнутые миомеры (рис. 273). При этом количество миомеров соответствует числу позвонков.
Среди мышц туловища лучше всех развита большая боковая мышца, которая продольным тяжом простирается от головы до хвоста, разделяясь посередине продольной соединительнотканной прослойкой - горизонтальной перегородкой. Над большой боковой мышцей находится прямая боковая поверхностная мышца (у лососевых в ней накапливается много жира), а по нижней стороне тела - прямая брюшная мышца (у некоторых, например у угрей, она отсутствует). Между последними двумя мышцами располагаются косые мышцы.

Рис. 273. Мускулатура костистой рыбы (окуня):
1 - мускулатура челюстного аппарата; 2 - миомеры; 3 - миосепты (по Кузнецову и соавт., с изменениями и дополнениями)
Поскольку основная масса мышечной ткани находится на спинной стороне тела, именно там у рыб располагается центр тяжести. Если у рыбы нарушается деятельность плавников или их попросту отрезать, рыба непременно переворачивается брюшной стороной вверх. Основной функцией мышц туловища является волнообразное изгибание тела в сагиттальной плоскости, что обеспечивает движение рыбы.
Мышцы головы управляют движениями челюстного аппарата и жаберных крышек, поэтому их еще называют висцеральной мускулатурой. Мускулатура плавников изменяет положение парных плавников и стабилизирует непарные плавники. В последнем случае по бокам от радиалий симметрично располагаются мышечные пучки. Эти мышцы не сегментированы.
Вся соматическая мускулатура образована поперечнополосатыми мышечными волокнами, которые развиваются непосредственно из миотомов сомитов. У рыб эти волокна уже дифференцированы на красные и белые. Напоминаем, что красные мышечные волокна содержат больше миоглобина (дыхательного пигмента, близкого к гемоглобину, также способного связывать молекулярный кислород), но меньше сократимых миофибрилл, кроме того, в саркоплазме много капель жира. Поэтому они способны к длительной работе, но не проявляют большой мощности. Эти волокна располагаются в мышцах более поверхностно. Белые волокна беднее миоглобином, но содержат гораздо больше сократительных миофибрилл, благодаря чему они способны мощно сокращаться, однако не так долго, как красные. Поэтому при медленном движении с постоянной скоростью сокращаются, в основном, красные волокна, тогда как резкое ускорение обеспечивают белые волокна. Основная часть мышечных волокон белая (например, у жереха они составляют 96,3%), однако у мигрирующих рыб доля красных волокон несколько больше.
Состояние мышечной системы зависит от многих факторов (возраста, образа жизни, величины и т.д.), например, у кеты (представителя лососевых рыб) после нереста мышечная ткань теряет почти весь жир (до 98,4%) и более половины (57%) белка.
Скопления специализированных мышечных волокон образуют элекрические органы рыб, которые могут располагаться в разных частях тела в зависимости от видовой принадлежности рыбы (чаще всего они находятся по бокам тела). Электрический орган напоминает аккумуляторную батарею, он состоит из особых мышечных пластинок, разделенных студенистой тканью. Чем мощнее электрический орган, тем больше в его составе пластинок.
Следует отметить, что слабые электрические разряды способны испускать многие виды, не имеющие специализированных органов (например, ставрида, окунь и др.). В морской воде такие разряды распространяются на 10 - 15 м (в пресной только на 2 м, поскольку в ней мало электролитов) и позволяют рыбе ориентироваться или посылать сигналы другим особям. Слабоэлектрические рыбы имеют специализированные электрические органы, правда, небольшие. Они способны испускать разряды менее 17 В, что позволяет хорошо ориентироваться в мутных водах. Наконец существуют виды, которые способны генерировать разряды до 600 В (например, электрический угорь, электрический сом). Столь мощные разряды способны временно оглушить жертву или врага и даже убить его. Неосторожное обращение с такими рыбами очень опасно для человека.
Пищеварительная система костных рыб включает в себя те же отделы, что и у хрящевых. Положение ротового отверстия зависит от способа питания рыбы. У планк- тоноядных форм рот занимает верхнее положение, у хищников он конечный, а у бентосоядных (подбирающих пищу со дна) - нижний, кроме того, встречаются различные переходные варианты (рис. 274). Некоторые виды имеют выдвижной рот, что позволяет им более эффективно рыться в илистом дне в поисках пищи (рис. 275).

Рис. 274. Положение рта у карповых рыб:
А - верхний у чехони: б - конечный у сазана; В - полунижний у воблы; Г - нижний у остролучки (по Никольскому)
Ротовая полость снабжена многочисленными зубами, которые несут целый ряд костей обоих отделов черепа (мозгового и висцерального): верхнечелюстные, межчелюстные, зубные кости (напоминаем, что все эти кости принадлежат челюстному аппарату), нёбные, сошник и копула подъязычной дуги. Все зубы обычно однотипны и наклонены по направлению к глотке, поэтому рыбы могут только удерживать добычу, или отрывать от нее куски (например, пираньи), но не могут ее жевать. Обычно зубы прирастают к поверхности кости, иногда они присоединяются подвижно. Основу зуба составляет дентин, покрытый снаружи тонким слоем эмали. В течение жизни изношенные зубы заменяются новыми. У мирных рыб (так называются виды, питающиеся растительной пищей или планктоном) в ротовой полости зубы отсутствуют. Так же, как и у хрящевых рыб, у костных отсутствует настоящий язык, имеющаяся вместо него складка слизистой оболочки лишена собственных мышц и поэтому не способна выдвигаться (у некоторых видов на этой складке могут даже находиться зубы).

Рис. 275. Выдвижной рот у сазана (по Никольскому)
Пронизанная жаберными щелями глотка активно участвует в процессах питания. Поступление пищи в ротовую полость происходит за счет движения жаберных крышек, при этом вода вместе с пищей засасывается в ротовую полость, из нее проходит в глотку и выталкивается из нее через жаберные щели. При этом пища остается в глотке, поскольку ее не пропускают жаберные тычинки, расположенные на жаберных дугах.

Рис. 276. Жаберные тычинки рыб:
А - ппанктоноядной; Б - бентосоядной; В - хищной: 1 - жаберные дуги; 2 - жаберные тычинки: 3 - жаберные лепестки (по И. М. Анисимовой и соавт., с изменениями и дополнениями)
Строение и количество тычинок у разных видов неодинаково и зависит от характера питания. У рыб, питающихся планктоном (планктонофагов), длинные многочисленные тычинки в совокупности образуют фильтрационный аппарат, отделяющий пищу от проходящей через жаберные щели воды (рис. 276). У видов, питающихся крупной пищей (хищных рыб), тычинки значительно короче или вообще не развиты, поскольку им нужно не отцеживать пищу, а удерживать ее. С этой задачей вполне справляются сами жаберные дуги, не пропуская крупные куски в жаберные щели. У некоторых видов на задней жаберной дуге имеются широкие и массивные глоточные зубы, которые перетирают пищу (рис. 277). Такие зубы обычно образуются у бентосояд- ных мирных рыб (например, у карповых), у которых в ротовой полости нет зубов. Слюнные железы у костных рыб, так же как и у хрящевых, отсутствуют.
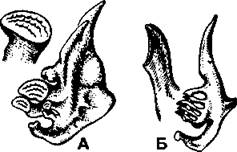
Рис. 277. Глоточные зубы карповых рыб: А - сазан; Б - лещ (по Никольскому)
Короткий пищевод ведет в желудок, который анатомически выражен не у всех видов, например, он отсутствует у карповых, некоторых бычков и др. Желудок переходит в тонкую кишку, в начальный отдел которой впадают протоки печени и поджелудочной железы. Именно в этом отделе кишечника расщепляется большая часть высокомолекулярных соединений пищевой кашицы, а также осуществляется всасывание. Тонкая кишка костных рыб значительно длинней, чем у хрящевых, она образует петли (первая из них называется двенадцатиперстной кишкой), что увеличивает всасывательную поверхность. У многих видов на границе между желудком и тонкой кишкой имеются пилорические придатки, в которых расщепляются белки и всасываются аминокислоты. Гистологически придатки соответствуют начальному отделу кишки, они обеспечивают значительное увеличение функциональной поверхности. Так, например, у радужной форели общая длина придатков превышает длину кишки в шесть раз, а поверхность всасывания превышает площадь внутренней поверхности тонкой кишки в 3,2 раза. Количество пилорических придатков варьирует от 3 (у окуня) до 400 (у лососей).
Печень крупная, обычно многолопастная (например, у карповых число лопастей может достигать семи), относительная масса печени костных рыб меньше, чем хрящевых (1 - 8% от массы тела), но функции ее те же. Желчный пузырь имеется. Анатомически оформленная поджелудочная железа отсутствует, ее структуры диссоциированы в ткани печени. Она выполняет экзокринную и эндокринную функции. Как экзокринная железа, она синтезирует и выделяет пищеварительные ферменты - трипсин, амилазу, мальтазу, липазу и др., которые через протоки поступают в полость тонкой кишки. В качестве эндокринной железы печень синтезирует некоторые гормоны, поступающие в кровь. У высших костных рыб (представителей надотряда костистых) эндокринные клетки уже обособляются в виде островков Лангерганса.
Тонкая кишка переходит в толстую, которая внешне практически не отличима от тонкой. Спиральный клапан имеется только у низших форм (осетровые, двоякодышащие), у высших костных рыб он отсутствует. Общая длина пищеварительного тракта широко варьирует у разных видов, что, прежде всего, связано с характером пищи - у растительноядных она длиннее (например, у толстолобика превышает длину тела в 6 - 15 раз), у всеядных рыб короче (например, кишечник карася длиннее тела в 2 - 3 раза), а у хищных рыб - совсем короткий (например, у судака или щуки составляет всего 0,6 - 1,2 длины тела).
Клоака имеется только у двоякодышащих рыб, у всех остальных кишечник открывается наружу анальным отверстием.
У большинства костных рыб имеется плавательный пузырь, который эмбрионально развивается как вырост дорзальной стенки передней части пищеварительного тракта. В зависимости от того, сохраняется ли связь пузыря с пищеводом, рыбы могут быть открытопузырными и закрытопузырными. У некоторых видов пузырь посредством группы мелких костей связан с перепончатым лабиринтом внутреннего уха, что позволяет рыбе воспринимать импульсы от изменяющего объем плавательного пузыря при смене положения тела в пространстве (рис. 278). Сообщение пузыря с органом равновесия называется аппаратом Вебера. Плавательный пузырь заполнен воздухом, что существенно уменьшает удельный вес рыбы и позволяет ей зависать в воде (напомним, что хрящевые рыбы, у которых плавательный пузырь отсутствует, прекращая движение, сразу же начинают погружаться в глубину). Рыбы могут произвольно изменять объем газа в пузыре, что ведет к уменьшению или повышению плавучести. У открытопузырных это происходит легко - путем обычного сжатия или расширения пузыря. Закрытопузырные рыбы для этого имеют в стенке пузыря специальную сеть кровеносных капилляров, которые поглощают или выделяют газ в зависимости от потребностей рыбы.
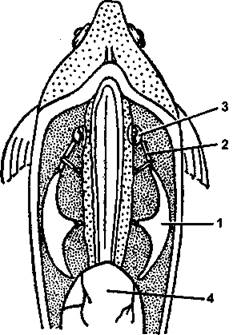
Рис. 278. Схема строения аппарата Вебера: 1 - рычаг: 2 - промежуточная кость; 3 - покрышка; 4 - плавательный пузырь
Дыхательная система, как и у всех первичноводных животных, представлена эктодермальными жабрами. Организация жаберного аппарата во многом сходна с аппаратом хрящевых рыб, но имеется ряд особенностей. Прежде всего частично (у осетровых) или полностью (у высших костных) редуцируются межжаберные перегородки, поэтому жаберные лепестки у них расположены непосредственно на жаберных дугах и омываются водой не с одной, а с обеих сторон (рис. 279, 280). Кроме того, у высших костных рыб отсутствует полужабра на подъязычной дуге, поэтому у них функционируют четыре пары жабр. Лишь у костных рыб имеется жаберная крышка, которая активно участвует в дыхательном акте и позволяет интенсифицировать газообмен при неподвижном положении рыбы. Дыхательный акт костных рыб включает в себя два этапа. На первом из них рыба приподнимает жаберные крышки, что создает в околожаберном пространстве пониженное давление и обеспечивает поступление свежей воды через ротовое отверстие. В щель между жаберными крышками и телом вода при этом не поступает, потому что эта щель оказывается прикрытой тонкой мембраной, которая продолжается за края жаберной крышки и при ее движении присасывается к телу, закрывая собой щель. На втором этапе отодвинутые жаберные крышки с силой прижимаются к телу, и возросшее в результате этого давление в околожаберной полости выталкивает оттуда воду во внешнюю среду. При этом вода не выходит изо рта, поскольку ротовое отверстие в тот момент закрывается (рис. 281).
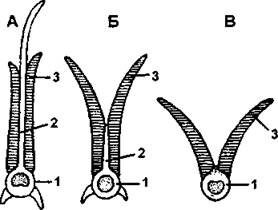
Рис. 279. Схема прикрепления жаберных лепестков у разных групп рыб:
А - хрящевые рыбы; Б - низшие костные (осетровые); В - костистые; 1 - жаберная дуга: 2 - межжаберная перегородка; 3 - жаберные лепестки (по С. И. Левушкину и соавт.)
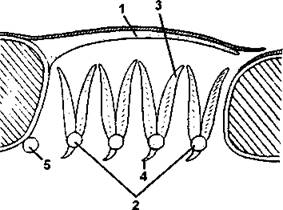
Рис. 280. Схема жаберного аппарата костистой рыбы:
1 - жаберная крышка; 2 - жаберные дуги; 3 - жаберные лепестки; 4 - жаберные тычинки; 5 - подъязычная дуга (по С. И. Левушкину и соавт.)
Такой способ поступления и удаления воды используется в состоянии покоя и при медленном движении, однако при высокой скорости плавания костные рыбы вполне могут переключаться на напорный тип вентиляции, описанный выше у акул, при котором вода пассивно поступает через открытое ротовое отверстие и омывает жабры. При этом существенно снижаются энергозатраты (до 30%), так как сокращается объем работы висцеральной мускулатуры и повышается эффективность движения (поскольку жаберные крышки при этом не создают дополнительных выступающих частей, снижающих обтекаемость тела рыбы).
Для рыб характерна очень высокая эффективность газообмена, например, некоторые виды высших костных рыб способны усваивать до 85% растворенного в воде кислорода, тогда как у хрящевых рыб этот показатель существенно ниже (70 - 75%). Это свойство чрезвычайно важно для обитателей водной среды, которая значительно беднее кислородом, нежели воздух (в среднем вода природных водоемов содержит около 5-11 мл/л растворенного О2). Для лучшего усвоения растворенного кислорода жаберные лепестки костных рыб имеют сложную структуру. На них расположены многочисленные вторичные жаберные пластинки, которые лежат перпендикулярно оси лепестка (рис. 282). Кроме того, эффективность газообмена возрастает из-за того, что движение воды, омывающей жабры, и крови в жаберных капиллярах осуществляется в противоположных направлениях (см. рис. 282). Поэтому вода уносит большую часть углекислого газа, диффундирующего из крови, и отдает в кровь большую часть кислорода, при этом выходящая из жаберной полости вода имеет почти такое же содержание этих газов, что и оттекающая от жабр кровь.

Рис. 281. Механизм дыхания взрослой рыбы: А - вдох; Б - выдох (по Никольскому)

Рис. 282. Схема противотока воды и крови в жабрах костистых рыб:
А - фрагмент жаберного лепестка с жаберными пластинками: 1 - артерия; 2 - вена: 3 - жаберная пластинка: 4 - кровь; 5 - жаберный лепесток; 6 - вода; Б - принципиальная схема противоточного обменника (цифрами обозначена степень насыщения газами воды и крови) (по Шмидт-Ниельсену и Ноак, с изменениями и дополнениями)
Столь совершенные механизмы газообмена приводят к тому, что рыба может использовать не все ресурсы дыхательной системы, например, установлено, что при достаточном насыщении воды кислородом кровоснабжаются только 60% вторичных жаберных пластинок. Однако кровоснабжение усиливается при повышении функциональных нагрузок или снижении содержания кислорода в воде. Общая поверхность газообмена у рыб достаточно велика и зависит от уровня активности рыб, в зависимости от этого величина показателя у разных видов широко варьирует (табл. 6).
Таблица 6
Дыхательная поверхность жабр (по Строганову)
|
Виды рыб |
Масса, г |
Дыхательная поверхность жабр |
|
|
см2 |
см2/кг |
||
|
Серебряный карась |
10,0 |
16,96 |
1700 |
|
Камбала |
135,0 |
889,00 |
6762,9 |
|
Окунь |
73,0 |
1173,8 |
16752,1 |
Рыбы, обитающие в условиях постоянного дефицита кислорода, выработали разнообразные адаптации. Например, при гипоксии учащаются дыхательные движения, увеличивается содержание в крови эритроцитов, что позволяет связывать кислород при его небольших парциальных давлениях.
Большое значение имеет кожный газообмен, причем его относительная доля зависит от условий, в которых находится рыба. Например, виды, постоянно обитающие в хорошо аэрируемых водах (например, сиги), не испытывают дефицита кислорода, поэтому у них доля кожного дыхания не превышает 3-9% от общего газообмена. Напротив, рыбы, которые живут в условиях регулярного дефицита кислорода (например, вьюны, сомы, угри, карпы и др.), имеют гораздо более высокую долю кожного газообмена (например, у вьюна через кожу поглощается до 85% кислорода и выделяется до 92% углекислого газа). Такие рыбы могут выдерживать длительное время вне воды и сохраняют жизнеспособность, например, во влажной среде карась способен жить до 11 суток, тогда как лещ гибнет через несколько часов.
Многие виды располагают дополнительными органами газообмена, с помощью которых усваивают кислород атмосферного воздуха. В наибольшей степени это выражено у двоякодышащих рыб и африканского многопера, плавательный пузырь которых видоизменился в ячеистое легкое, густо оплетенное кровеносными сосудами. У этих рыб даже имеется второй - «легочный» круг кровообращения. При этом используется не только кислород воды (при жаберном дыхании), но и кислород атмосферного воздуха (при легочном дыхании). Воздушный газообмен очень эффективен - при легочном дыхании кровь насыщается кислородом на 90%. В меньшей степени плавательный пузырь служит газообмену и многим другим рыбам (например, панцирной щуке, ильной рыбе и др.).
У лабиринтовых рыб в качестве дополнительного органа газообмена образуется кармановидное расширение участка жаберной полости - лабиринт. В этом месте имеются складчатые стенки, обильно пронизанные сосудами, в которых усваивается кислород воздуха. К лабиринтовым рыбам относятся известные аквариумистам макроподы, гурами, лялиусы, петушки и др. У змееголова воздушный газообмен осуществляется в сосудах наджаберной полости, которая представляет собой вырост глотки.
Менее выраженные адаптации к воздушному газообмену имеются у многих рыб. Например, при недостатке кислорода (например, в покрытом длительное время ледяным слоем водоеме) рыбы скапливаются у проруби и захватывают воздух, который насыщает кислородом воду, находящуюся в ротовой полости. Часто рыбы глотают воздух (в прямом, а не переносном смысле), после этого кислород всасывается капиллярами, которые оплетают стенки кишечника. Такой способ газообмена, свойственный только рыбам, довольно продуктивен (с его помощью из воздуха усваивается до 50% кислорода).
Однако избыток кислорода также губителен, как и его недостаток. Установлено, что перенасыщение воды растворенным кислородом приводит к гибели многих рыб (например, линя, карася и др.). При этом нарушаются акты дыхания и координация движений, жабры покрываются пузырьками, кровь становится пенистой. Слишком большое содержание кислорода приводит к нарушениям инкубации икры, при этом отмечается значительная гибель икринок, а среди молоди много уродов.
Кроме кислорода, на ход газообмена оказывает влияние углекислый газ. При чрезмерном содержании этого газа в воде гемоглобин в притекающей к жабрам венозной крови не теряет связи с углекислым газом и поэтому не связывается с кислородом. Установлено, что содержание углекислого газа в атмосферном воздухе свыше 1 — 5% также делает невозможным связывание гемоглобином растворенного кислорода, и рыба гибнет.
Кровеносная система во многом сходна с хрящевыми рыбами (рис. 283), поэтому мы не будем ее рассматривать целиком, а остановимся лишь на особенностях.
Артериальный конус, свойственный желудочку хрящевых рыб, имеется только у низших костных (осетровых, кистеперых, двоякодышащих), у более высокоорганизованных вместо него образуется луковица аорты (рис. 284), которая является расширением начального отдела брюшной аорты и к желудочку не относится (доказательством этому служат гладкие миоциты стенки луковицы), поэтому самостоятельно пульсировать не может.
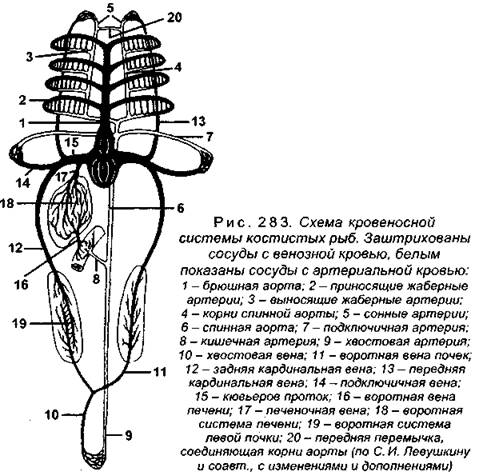

Рис. 284. Строение сердца костистых рыб:
1 - венозный синус: 2 - предсердие; 3 - желудочко-аортальный клапан; 4 - брюшная аорта; 5 - луковица аорты; 6 - желудочек; 7 - предсердно-желудочковый клапан (по С. И. Левушкину и соавт., с изменениями и дополнениями)
Артериальная система характеризуется редукцией передней пары жаберных артерий (приносящих и выносящих), поскольку на подъязычной дуге отсутствует полужабра (имеющаяся у хрящевых рыб). Таким образом, сохраняются лишь четыре пары приносящих и выносящих жаберных артерий.
Парные корни спинной аорты соединяются сзади, формируя спинную аорту, и спереди, образуя характерный для высших костных рыб артериальный головной круг.
Венозная система отличается тем, что отсутствуют боковые вены, кроме того, у большинства видов только левая задняя кардинальная вена образует воротную систему в почке, а правая идет не прерываясь (см. рис. 283).
Значительные изменения в организации кровеносной системы имеются у рыб, освоивших дыхание атмосферным воздухом, при этом у двоякодышащих рыб даже образуется второй круг кровообращения.
Форменные элементы крови у костных рыб относятся к тем же группам, что и у хрящевых. Наиболее многочисленными являются эритроциты, количество которых у разных видов широко варьирует и зависит, прежде всего, от степени подвижности рыбы (например, у малоподвижного карпа их примерно 0,84 млн./мм3 крови, тогда как у пеламиды - 4,12 млн./мм3). Лейкоцитов значительно меньше, они бывают зернистыми (в основном, нейтрофилы) и незернистыми (в основном, лимфоциты).
У большинства видов дыхательный пигмент (гемоглобин) находится в эритроцитах, при этом относительное содержание гемоглобина невелико (примерно 0,5-4 г/кг массы тела, тогда как у млекопитающих 5-25 г/кг). Количество гемоглобина не постоянно и зависит от сезона, например, у карпа зимой выше, чем летом, что можно считать адаптацией к гипоксии, поскольку зимой насыщенность воды кислородом снижается. У некоторых представителей подотряда нототениевых (отряд окунеобразных), обитающих в богатых кислородом холодных водах Антарктики, гемоглобин отсутствует и кислород растворяется непосредственно в плазме крови, кроме того, у них наряду с жаберным большое значение имеет кожный газообмен (на 1 мм2 поверхности кожи приходится до 45 мм капилляров).
Органы кроветворения такие же, как и у хрящевых рыб, но у высших костных рыб наиболее активно гемопоэз идет в почках, а также в лимфоидных органах и селезенке.
Лимфатическая система костных рыб развита значительно лучше, чем у хрящевых. Имеется выраженная система сосудов, по которым лимфа оттекает от внутренних органов и тканей. В конечном итоге лимфатические сосуды впадают в вены (обычно в кювьеровы протоки).
Выделительная система, как и у хрящевых рыб, представлена лентовидными мезонефрическими почками, расположенными под брюшиной по бокам позвоночного столба над плавательным пузырем. Мочеточники соединяются в выводной канал, который открывается не в клоаку, а на конце мочеполового сосочка. Передний отдел почек представляет собой сильно редуцированный, но сохранившийся пронефрос. У взрослых рыб он составлен лимфоидной тканью и выполняет функцию кроветворного органа, также в нем задерживаются и погибают старые эритроциты. Почка быстро реагирует на состояние рыбы (например, при недостатке в воде кислорода она уменьшается в объеме).
У примитивных костных рыб (осетровых) в нефронах сохраняется воронка (нефростом), однако у более высокоорганизованных форм она отсутствует. У хрящевых рыб концентрация солей в жидкостях внутренней среды организма ненамного отличается от аналогичного показателя морской воды (подавляющее большинство хрящевых рыб - обитатели моря), поэтому им несложно поддерживать водно-солевой баланс. Внутренняя среда костных рыб содержит иное количество солей, чем в окружающей воде (у пресноводных форм в крови солей больше, а у морских - меньше), поэтому они вынуждены искусственно поддерживать водно-солевое равновесие внутренней среды.
У пресноводных рыб кровь гипертонична по отношению к внешней воде, поэтому, хоть активного питья не происходит, внутрь тела за счет разницы осмотического давления постоянно проникает большое количество излишней воды (через жабры, кожу, ротовую полость, слизистую кишечника). Как мы помним, в сходном положении оказываются пресноводные простейшие (см. раздел, посвященный подцарству простейшие), которые решили проблему выведения излишней воды с помощью сократительной вакуоли, присущей только им. Пресноводные костные рыбы удаляют воду с помощью сильно развитых почек с многочисленными нефронами, которые выделяют огромное количество (более 300 мл/кг массы тела в сутки) слабоконцентрированной (гипотоничной) мочи. Дефицит солей при этом компенсируется активным всасыванием ионов из окружающей воды с помощью жабр, а также реабсорбцией ионов из первичной мочи в канальцах мезонефроса. Главным конечным продуктом азотистого обмена у них является не мочевина (как у хрящевых рыб), а аммиак, который значительно токсичнее и требует гораздо большего разбавления, поэтому большие объемы выделяемой мочи оказываются весьма кстати.
Между тем пресноводные рыбы могут жить и в соленых водоемах, удаляя излишние соли (они поступают с пищей и путем диффузии через жабры) с мочой, фекалиями и частично через кожу.
Осморегуляция у морских рыб осуществляется иначе. У них кровь гипотонична по отношению к внешней воде, поэтому перед ними стоят противоположные проблемы - им нужно удалять лишние соли и удерживать воду, которая теряется осмотическим путем (главным образом, через жабры). Понятно, что выделять так много мочи, как это делают пресноводные рыбы, морским нельзя, поэтому у них в почках меньше нефронов, размеры клубочков малы (у некоторых они даже могут вообще отсутствовать, например у семейства Syngnathidae и некоторых антарктических рыб), причем значительная часть их вообще не функционирует (не участвует в фильтрации). Сохранение не действующих клубочков полезно, поскольку соленость воды может изменяться и эти структуры, возможно, понадобятся в будущем. При этом значительно снижается объем выделяемой мочи, например, бычок выделяет всего 3-23 мл/кг в сутки. Аналогично ведут себя пресноводные рыбы, оказавшись в морской воде, например, угорь в пресной воде образует 60 - 150 мл/кг в сутки, а в морской - 2 — 4 мл/кг в сутки (по данным Строганова, 1962). Легко догадаться, что ограничение потерь воды с мочой не снимает проблему дефицита воды, а только ее облегчает, поскольку воду необходимо где-то брать. Поэтому морские рыбы попросту воду пьют, чего не делают их пресноводные сородичи. Поступившие с морской водой соли удаляются затем через почки, где ионы не задерживаются (как у пресноводных форм), а выводятся, тогда как вода подвергается реабсорбции и поступает в сосудистое русло. Кроме того, всасывание воды происходит не только в почках, но и в мочевом пузыре (он имеется у некоторых видов). Соли выделяются также через жабры. У некоторых проходных рыб в осморегуляции участвует выделяемая кожей слизь.
Половая система костных рыб очень своеобразна и совершенно отлична от хрящевых рыб. Мужская половая система состоит из парных вытянутых мешкообразных семенников, от стенок которых внутрь органа отходят многочисленные семенные канальцы, объединяющиеся по направлению к выводному протоку. Если канальцы сильно извиваются в различных плоскостях, то семенник относят к цип- риноидному типу (рис. 285), такие семенники обычно имеют округлые края, а выводной проток располагается в верхней части органа. Семенники перкоидного типа характеризуются радиальным ходом семенных канальцев, при этом выводной проток находится в центре органа, который на поперечном срезе имеет треугольную форму. Циприноид- ные семенники имеются у осетровых, карповых, сомовых, щуковых, тресковых, сельдевых и др., а перкоид- ные — у колюшковых, окуневых и др.

Рис. 285. Типы строения семенников костистых рыб: А - перкоидный: Б - циприноидный (по И. М. Анисимовой и совет.)
В отличие от хрящевых, семявыносящие протоки семенников костных рыб самостоятельные, они открываются общим половым отверстием позади анального отверстия. Таким образом, мужские половые протоки костных рыб не имеют никакого отношения к вольфовым протокам, которые у них служат только мочеточниками.
Женская половая система представлена парными (у самок некоторых видов, например у окуня, яичник одиночный) удлиненными мешкообразными полыми яичниками, занимающими в теле примерно такое же положение, что и семенники. Часть оболочки яичника вытягивается и образует проток, через который созревшие яйца (их у рыб обычно называют икринками) выходят либо в мочеполовой синус, либо прямо наружу через самостоятельное непарное половое отверстие, лежащее позади анального отверстия (рис. 286). Таким образом, у самок костных рыб мюллеровы протоки редуцируются. У самок разных видов костных рыб могут иметься различные особенности в организации половой системы.
Развитие. У большинства костных рыб оплодотворение внешнее, при этом процесс откладывания яиц (икры) называется нерестом. У некоторых видов он может сопровождаться сложным поведением половых партнеров, особенно сложные ритуалы демонстрируют различные виды цихлид, причем эти рыбы отличаются тем, что выбирают себе партнера (иногда на всю жизнь) в соответствии с собственными взглядами, например, в аквариуме их невозможно принудить вывести потомство с «несимпатичным» представителем противоположного пола. Часто во время нереста изменяется строение рыбы. В отличие от хрящевых, многие виды костных рыб проявляют заботу о потомстве. Это относится к тем же цихлидам, у которых потомство охраняют оба родителя. Самцы лабиринтовых рыб (например, гурами) из пены строят плавучее гнездо, куда помещают икринки (интересно, что самки после икрометания изгоняются самцом и о потомстве не заботятся). Гнездо из травинок строит самец трехиглой колюшки, затем несколько дней присматривает за выклюнувшейся молодью. Интересно заботится о потомстве африканская тиляпия - рыбка вынашивает икринки во рту, но и после выхода молоди в случае опасности мальки быстро заплывают в большой рот родителя и находятся там, пока обстановка не станет спокойной.
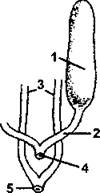
Рис. 286. Мочеполовой аппарат самок костистых рыб: 1 - яичник; 2 - яйцевод: 3 - мочеточник; 4 - половые отверстия; 5 - мочевое отверстие (по Шмальгаузену)
Многие виды для нереста мигрируют из моря в реки, обычно туда, где когда-то сами появились на свет (такие рыбы называются проходными). Они демонстрируют удивительные навигационные способности, когда безошибочно находят дорогу за тысячи километров к нужной реке, выбирая ее среди множества других. Полагают, что здесь помогает тонкое обоняние, поскольку лососи сильно возбуждаются, если в окружающую воду добавить воду из родной реки, но остаются невозмутимыми при добавлении воды из реки «чужой». Европейские угри, напротив, во взрослом состоянии живут в реках Европы, но для нереста совершают длительную миграцию в Саргассово море, где на глубине около 200 м нерестятся.
Количество икринок, которое мечет самка, широко варьирует от единичных до астрономических (например, самка луны-рыбы мечет до 30 млн. икринок - рекорд даже среди рыб). Икра некоторых видов свободно плавает в воде, других прикрепляется к водным растениям, поверхности донных камней и других подводных предметов, при этом кладка икры у разных видов выглядит неодинаково. Икринки обычно имеют мелкие размеры, покрыты тонкой оболочкой и содержат много желтка, расположенного вблизи одного из полюсов (тело- лецитальные яйца). Развитие костных рыб чаще всего идет с прохождением личиночной стадии (рис. 287).

Рис. 287. Развитие молоди карпа после выклева:
А - только что выклюнувшаяся првдличинка, L = 5.0 мм; Б - личинка, L = 7,5 мм (3-й сутки); В - личинка, L = 8,5 мм (4-е сутки); Г -личинка, L = 12,5 мм (8-е сутки); Д - личинка, L = 16 мм (14-е сутки) (по И. М. Анисимовой и соавт.)
Среди костных рыб имеются и живородящие формы, к ним относятся, например, хорошо известные аквариумистам гуппи, меченосцы, моллиенезии и некоторые другие, при этом количество рожденных детенышей может быть достаточно большим, например, бельдюга рождает 100 - 300 детенышей. Значительная часть икринок поедается другими рыбами, как правило, гибнет много молоди, поэтому до взрослого состояния доживает лишь незначительная часть потомства. При этом замечено, что у менее плодовитых рыб относительная доля выжившей молоди значительно выше, чем у более плодовитых.
Нервная система имеет то же разделение, что и у хрящевых рыб. Головной мозг устроен во многом примитивнее, чем у хрящевых рыб (рис. 288). Относительные размеры мозга очень малы (0,02 - 0,94% от массы тела), конечный мозг меньше, чем у хрящевых рыб, он не делится даже неполной перегородкой, поэтому его полость едина и не образует боковых желудочков. Нервные элементы содержатся только в основании и боковых стенках конечного мозга (его крыша образована эпителием). Нижний придаток промежуточного мозга - гипофиз - состоит из передней и промежуточной долей. Интегративным центром является относительно большой средний мозг, поэтому головной мозг костных рыб относят к ихтиопсидному типу. Мозжечок тоже имеет значительные размеры, особенно у подвижных видов (например, пелагических рыб или активных хищников). У малоподвижных рыб (например, камбал) мозжечок мал, но относительно хорошо развиты конечный и продолговатый мозг, что особенно важно для восприятия растворенных в воде веществ (обоняние) и осязания. Рыбы не так быстро формируют условные рефлексы, как это делают высшие позвоночные, и быстрее их теряют, но в течение жизни они не только способны воспринимать изменяющиеся условия внешней среды, но и оптимально адаптировать к ней свое поведение.
Организация спинного мозга не имеет существенных отличий от хрящевых рыб.

Рис. 288. Головной мозг окуня:
1 - обонятельная капсула; 2 - обонятельные доли; 3 - передний мозг; 4 - средний мозг; 5 – мозжечок; 6 - продолговатый мозг; 7 - спинной мозг; 8 - глазничная ветвь тройничного нерва; 9 - слуховой нерв; 10 - блуждающий нерв (по Суворову)
Вегетативная нервная система организована более сложно, чем у хрящевых рыб. Расположенные вблизи позвоночного столба ганглии не разобщены, а соединяются в два продольных симпатических ствола (правый и левый). Белые соединительные ветви соединяют вегетативные стволы с ЦНС, а серые соединительные ветви - с иннервируемыми органами. Парасимпатический отдел представлен вегетативными волокнами блуждающего нерва.
Органы чувств в основном те же, что и у хрящевых рыб. Орган зрения имеет обычное для рыб строение (рис. 289), плоская роговица и почти шаровидный хрусталик более эффективны в водной среде, но позволяют отчетливо различать только ближние предметы. От сосудистой оболочки глаза вблизи зрительного нерва отходит серповидный отросток, который вдается в полость глазного яблока и прикрепляется к хрусталику. Когда этот отросток сокращается, он отодвигает хрусталик в глубь глаза, обеспечивая тем самым аккомодацию. Кроме того, характерна серебристая оболочка, которая представляет собой слой сосудистой оболочки, богатый отложениями кристалликов гуанина. Эта оболочка располагается между сосудистой и фиброзонй (белочной) оболочками, впереди она переходит на радужку, образуя ее наружный слой, из-за чего глаз приобретает зеленовато-золотистый блеск.

Рис. 289. Глаз костистой рыбы, вертикальный разрез:
1 – хрусталик; 2 – роговица; 3 - радужка: 4 - сосудистая оболочка; 5 - пигментная оболочка; 6 - серебристая оболочка; 7 – сетчатка; 8 - склера с костными отложениями внутри; 9 - вздутие серповидного отростка; 10 - серповидный отросток; 11 - железа сосудистой оболочки; 12 - глазной нерв (по Паркеру)
Количество фоторецепторных клеток в сетчатке достаточно велико (например, у карпа их насчитывается около 50 000 на 1 мм2 сетчатки). Среди рецепторов различают палочки, которые реагируют на слабый свет, но не могут создать четкого изображения, и колбочки, нуждающиеся в большом количестве света. В зависимости от уровня освещения фоторецепторы располагаются в толще сетчатки по-разному. Если света много, колбочки перемещаются ближе к свету, а палочки оказываются прикрытыми расширившимися пигментными клетками. Соответственно, при слабом освещении все происходит наоборот - колбочки смещаются в глубь сетчатки, а палочки - на ее поверхность. Соотношение фоторецепторов зависит от образа жизни, например, у дневных рыб больше колбочек, а у ночных и глубоководных их практически нет, но многочисленны палочки. Например, у ночной рыбы налима палочек в сетчатке в 14 раз больше, чем у дневного хищника щуки, у некоторых глубоководных рыб количество палочек становится огромным — до 25 млн./мм2 сетчатки. Поскольку в сетчатке имеются колбочки, рыбы способны различать некоторые цвета (разумеется, те виды, которые обладают такими фоторецепторами).
У разных видов в строении глаза могут быть свои особенности, например, у кефали и у некоторых сельдей имеется мигательная перепонка. Глаза четырехглазки (обитает в Центральной и в Южной Америке) разделены на две половины - нижняя имеет плоскую роговицу, позволяющую хорошо ориентироваться в воде, а верхняя - выпуклую, которая обеспечивает зрение в воздушной среде. У некоторых видов зрение вторично редуцируется, такие виды ориентируются с помощью других органов чувств.
Орган слуха, как и у хрящевых рыб, представлен внутренним ухом, сочетающим в себе рецепторные поля двух анализаторов - слухового и вестибулярного (рис. 290). Строение внутреннего уха обычно для позвоночных - внутри костного лабиринта слуховой капсулы (его внутренние стенки не костные, а хрящевые!) находится перепончатый лабиринт, который повторяет форму костного и заполнен эндолимфой, в пространстве между двумя лабиринтами циркулирует перилимфа. В отличие от акул, эндолимфатический проток заканчивается слепо в коже головы. Наряду с мелкими отолитами в эндолимфе плавают крупные, которых у большинства высших рыб три, самый крупный из которых находится в круглом мешочке, заполняя его почти целиком.
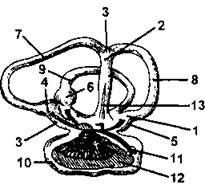
Рис. 290. Орган слуха окуня:
1 - овальный мешочек; 2 - верхний синус мешочка; 3 - верхушка мешочка; 4, 5 и 6 - ампулы полукружных каналов; 7, 8 и 9 - полукружные каналы; 10 - круглый мешочек; 11 - ветви слухового нерва; 12- отолит; 13 - эндолимфатический проток (по Суворову)
Рецепторы, воспринимающие звук, находятся в круглом мешочке. Низкочастотные звуки (от 5 до 25 Гц) рыбы воспринимают боковой линией, а высокочастотные (16 - 13 000 Гц) - внутренним ухом. Диапазон улавливаемых звуков у разных видов широко варьирует, например, угорь воспринимает звуки частотой 36 - 650 Гц, а язь - 25 - 5524 Гц. Следует отметить, что рыбы чувствуют звуки, которые генерируются в воздухе, несмотря на то, что в воду при этом попадает всего 0,01% звуковых волн.
Звуки играют большую роль в жизни рыб, позволяя ориентироваться при поиске корма, межвидовой коммуникации, брачном поведении и т.д. При этом они способны не только улавливать звуки, но и сами издают их (скрипы, щелчки, шорохи и т.п.). Источником звука могут быть различные структуры: плавательный пузырь, кости жаберной крышки, лучи грудных плавников, зубы (челюстные и глоточные) и др. Таким образом, несмотря на метафоричное «нем как рыба», рыбы вполне могут общаться с помощью звуков. Водная среда значительно усложняет механизм их образования, но скорость проведения звука в воде примерно в пять раз выше, чем в воздухе, и достигает 1500 м/сек. Особенно важна звуковая коммуникация у глубоководных рыб, которые постоянно находятся в условиях темноты.
Органы обоняния представлены обонятельными мешками, ноздри (передняя и задняя) расположены между ртом и глазами. Вода проникает через переднее отверстие, омывает сенсорный эпителий и вытекает через заднее отверстие. У быстроплавающих Арыб ноздри небольшие, тогда как у малоподвижных они очень крупные, что позволяет обеспечить достаточное поступление анализируемой воды к обонятельным рецепторам. У многих рыб обоняние очень тонкое, например, лососи чувствуют воду родной реки (в которой они появились на свет из икринки) на расстоянии 800 км и безошибочно находят дорогу к ней для нереста.
Органами вкуса являются мелкие вкусовые почки, которые разбросаны по всей наружной поверхности кожи, но особенно они многочисленны в слизистой оболочке ротовой полости, глотки, на жаберных лепестках, усиках, плавниковых лучах. Вкусовая почка образована скоплениями рецепторных клеток, среди которых располагаются опорные клетки. Чувствительные клетки оплетены нервными окончаниями лицевого, блуждающего и языкоглоточного нервов и снабжены на конце волоском. Установлено, что рыбы определяют сладкое, соленое, кислое и горькое.
Кроме того, хеморецепцию (восприятие химических раздражителей) осуществляют свободные окончания тройничного и блуждающего нервов, а также спинномозговых нервов.
Органы боковой линии имеют типичное строение, они представлены погруженным в кожу каналом, который тянется вдоль всего тела и сообщается с окружающей водой посредством многочисленных отверстий, пронизывающих чешуи. Вода свободно проникает через отверстия в канал, течет по нему и анализируется чувствительными клетками, которые этот канал выстилают. Группа рецепторных клеток вместе с окружающими их нервными волокнами составляют непосредственно чувствительный орган - невромаст.
Для многих видов рыб большое значение имеют органы, воспринимающие электрическое и магнитное поля. Такие органы расположены на всей поверхности тела, но больше всего их на голове и вблизи нее. Особенно важно восприятие электромагнитных сигналов для рыб, обитающих в мутной воде, где видимость практически отсутствует и ориентация с помощью зрения становится невозможной. Например, нильский слоник постоянно генерирует электромагнитные импульсы и воспринимает отраженные волны, анализируя полученную информацию, рыбка отлично ориентируется в непрозрачной воде.
Поведение костных рыб сложнее, чем у хрящевых, условные рефлексы многочисленны, причем вырабатываются значительно легче, а затухают медленнее. Это позволило костным рыбам отлично приспосабливаться к самым разнообразным условиям обитания, что нашло отражение в более высоком (чем у хрящевых рыб) видовом разнообразии.