Размножение лишайников столь же своеобразно, как и вся их организация. Каждый из компонентов таллома способен размножаться индивидуально, например, клетки водоросли время от времени делятся, однако последующее развитие нового таллома возможно лишь в случае повторной встречи гриба и водоросли. Поэтому более перспективным (во всяком случае, для микобионта) будет такой способ размножения, при котором генеративные структуры содержат оба компонента.
Большинство лишайников размножаются преимущественно вегетативным путем, причем для многих из них этот способ размножения остается единственным. Высыхая, слоевище становится очень хрупким и от него легко отламываются кусочки, которые затем подхватываются ветром и распространяются. Так происходит у многих лишайников тундры.
Соредии и изидии являются специализированными структурами вегетативного размножения, причем каждая из них содержит элементы микобионта и фикобионта. Соредии (греч. soros - куча) представляют собой микроскопические комочки клеток водоросли, окруженные гифами гриба (рис. 392). Скопление соредий (они называются соралями) напоминает порошкообразную массу, которая покрывает сверху слоевище или окружает таллом по внешнему периметру. Образование соредий возможно только в слое таллома, содержащем водоросли (гонидиальном слое), и через разрывы верхнего корового слоя соредии выходят наружу, образуя скопления - сорали. Соредиями размножаются главным образом высокоорганизованные лишайники (листоватые и кустистые), которые обычно не образуют плодовых тел.

Рис. 392. Вегетативное размножение лишайников с помощью соредий:
А - соредий (1 - клетки водорослей; 2 - гифы гриба);
Б - схема образования соредий в слоевище;
В-3- различная форма соралей лишайников:
В - пятновидная (Physcia orbicularis);
Г - головчатая (Hypogymnia bitten);
Д - манжетовидная (Menegazzia pertusa);
Е - зубовидная (Physcia tenella);
Ж-в виде каймы (Cetraria pinastri);
З - щелевидная (Parmelia sulcata)
(пo H. С. Голубковой)
Изидии также свойственны листоватым и кустистым формам, но они образуются реже, чем соредии. Морфологически изидии представляют собой мелкие выросты на верхней поверхности таллома (рис. 393), внутри которых находятся водорослевые клетки, окруженные гифами. Снаружи изидии покрыты коровым слоем. Внешний вид изидий может быть самым разнообразным, но для каждого вида лишайников специфична определенная форма.

Рис. 393. Вегетативное размножение лишайников с помощью изидий
А - слоевище листоватого лишайника ParmeHa exasperatula с изидиями (1);
Б - различная форма изидий (2 - шаровидная; 3 - булавовидная; 4 - цилиндрическая; 5 - коралловидная);
В - поперечный разрез через слоевище с изидиями (6- коровой слой; 7 - гифы гриба; 8 - водоросли) (по Н. С. Голубковой)
Бесполое размножение лишайников у каждого из компонентов происходит самостоятельно. Микобионт может образовывать конидии, пикноконидии и стилоспоры, формирующиеся экзогенно. Конидии, которыми размножаются многие свободноживущие грибы, у лишайников в природе образуются очень редко, гораздо чаще их можно наблюдать при искусственном культивировании микобионта. Пикноконидии представляют собой очень мелкие клетки (ширина около 1 мкм и длина около 5 мкм) различной формы, которые в огромном количестве образуются в замкнутых вместилищах округлой формы с отверстием в верхней части, называемых пикнидия- ми (греч. pyknos - плотный). Обычно они развиваются на верхней поверхности таллома или по его краям. Стилоспоры также развиваются в пикнидиях (только реже), но имеют большие размеры (до 100 мкм в длину) и состоят из двух и более клеток.
Половое размножение характерно для микобионта лишайников, оно сопровождается формированием плодовых тел. В зависимости от систематического положения микобионта различают базидиальные и сумчатые лишайники. Базидиальные лишайники (Basidiolichenes) в качестве микобионта имеют базидиальные грибы. Эта совсем небольшая группа (всего около 20 видов) имеет самостоятельное происхождение и распространена преимущественно в тропиках, хотя некоторые из них обитают в умеренном климате и даже в тундре.
Базидии у них образуются на нижней стороне слоевища в открытом генеративном слое, где базидии перемежаются со стерильными гифами - парафизами (рис. 394). Плодовые тела не тропических базидиаль- ных лишайников напоминают миниатюрные шляпочные грибы. Развитие базидиоспор во многом сходно с процессом полового спороношения у самостоятельных базидиальных грибов, о чем подробно рассказано в разделе настоящего пособия, посвященном этим грибам.

Рис. 394. Базидиальные лишайники:
А - тропический лишайник Cora pavonia;
Б-Г- базидиальные лишайники с плодовыми телами в форме шляпочного гриба (Б, В - Omphalina luteolilacina, внешний вид и поперечный разрез через плодовое тело;
Г - Omphalina ericetorum): 1 - базидии; 2 - парафизы; 3 - споры;
Д - роговидные плодовые тела базидиального лишайника Clavulinopsis septentrionalis (по Н. С. Голубковой)
Большинство лишайников содержат сумчатые грибы, поэтому они объединяются в группу сумчатых лишайников (Ascolichenes). При половом спороношении у них также образуются плодовые тела, развитие которых у разных форм происходит неодинаково. У асколокулярных лишайников вначале образуется строма, состоящая из переплетенных гиф, после чего в ней формируются женские половые органы - архикарпы. Развитие асков со спорами сопровождается образованием вокруг сумок полостей в строме - локул.
У большинства сумчатых лишайников развитие архикарпа начинается на границе сердцевины и слоя водорослей (гимениального слоя), поэтому у них в плодовых телах всегда имеется гимениальный слой. Процесс оплодотворения у лишайников изучен недостаточно, многие исследователи полагают, что плодовое тело развивается из неоплодотворенного аскогона.
У сумчатых лишайников выделяют следующие основные типы плодовых тел: апотеции, гастротеции и перитеции.
Апотеции (греч. apotheke - хранилище) являются плодовыми телами открытого типа, которые наиболее широко распространены среди лишайников. Они представляют собой мелкие (обычно 1-2 мм, реже крупнее или мельче) структуры округлой или овальной формы, расположенные на верхней (очень редко, только у нефромы и у нефромопсиса, на нижней) поверхности таллома. Апотеций состоит из уплощенной центральной части - диска и валика, окружающего этот диск по периферии. Репродуктивной функцией апотеция обладает диск, верхний слой которого (гимениальный слой) образован многочисленными сумками, ориентированными вертикально, и стерильными гифами (парафизами). Верхние концы парафиз имеют булавовидные утолщения, которые, смыкаясь, образуют над сумками (они лежат несколько ниже) защитный слой - эпитеций. Именно от эпитеция зависит окраска апотеция. Под гимениальным слоем находится слой плотно переплетенных гиф - гипотеций, в котором развиваются аскогенные гифы и находятся основания парафиз.
Валик, окружающий диск с боков и снизу, выполняет защитную функцию. В зависимости от строения валика, различают несколько типов апотециев: лекано- ровые, лецидиевые и лецидеевые.
Леканоровые апотеции характеризуются тем, что их диск окружен валиком, сходным по внутреннему строению и окраской с талломом, причем окраска валика и диска всегда разная. В составе валика (так же как и таллома) выделяются коровой слой, слой водорослей и более рыхлая сердцевина (рис. 395). Название этого типа плодовых тел происходит от латинского названия рода леканора.
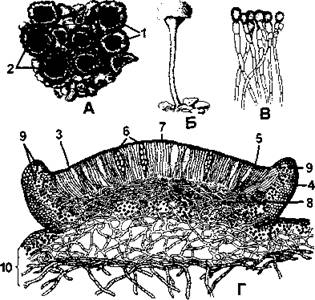
Рис. 395. Леканоровые апотеции:
А - внешний вид апотециев на накипном слоевище Lecanora ( 1 - диск; 2 - край);
Б - апотеции на длинной ножке; пылевидная масса наверху - мазедий;
В - парафизы с булавовидными окрашенными вершинами;
Г - поперечный разрез через апотеции (3 - гимениальный слой; 4 - слоевищный край; 5 - парафизы; 6 - сумки со спорами; 7 - эпитеций, 8 - гипотеций; 9 - водоросли, 10 - слоевище) (по Н. С. Голубковой)
Лецидеевые апотеции имеют валик, окрашенный так же, как и диск (обычно в черный цвет), при этом они отличаются высокой твердостью. Строение валика иное, чем основной таллом, его образуют плотно прилегающие друг к другу темноокрашенные гифы, которые в совокупности составляют эксципул, окружающий со всех сторон диск (рис. 396). Лецидеевые апотеции образованы только грибными гифами и не содержат водорослей. Название этого типа плодовых тел происходит от латинского названия рода лецидея.
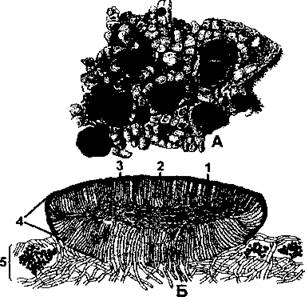
Рис. 396. Лецидеевые апотеции:
А - внешний вид темноокрашенных апотециев на накипном слоевище (Lecidea);
Б - поперечный разрез через апотеций (1 - парафизы; 2 - сумки со спорами; 3 - эпитеций; 4 - собственный край, образованный эксципулом; 5 - слоевище) (no Н. С. Голубковой)
Биаторовые лишайники во многом похожи на лецидеевые, но, в отличие от них, имеют более мягкую консистенцию и, кроме того, окрашены в светлые тона и никогда не бывают черными.
Гастеротеции (греч. gaster - желудок, theke - сумка) также являются открытыми плодовыми телами, но они встречаются очень редко (только у двух родов накипных лишайников: опеграфа и грифис). Их диск, располагающийся глубоко, имеет вид ; узкой полоски. Вокруг диска возвышается край, который может принадлежать слоевищу или самому гасте- ротецию (рис. 397).

Рис. 397. Гастеротеции (по Н. С. Голубковой)
Перитеции (грвч. peri - возле, около, theke - сумка), в отличие от апотециев и гастеротециев, являются плодовыми телами закрытого типа. Морфологически они представляют собой округлые или кувшинообразные структуры, глубоко погруженные в слоевище (рис. 398). Перитеции сообщаются с окружающей средой через маленькие отверстия на вершине, которые иногда называют устьицами (не путать с настоящими устьицами высших растений!). Через эти же отверстия зрелые споры покидают перитеций. В составе стенки плодового тела выделяют наружный слой (эксципул), образованный несколькими слоями гиф, иногда снаружи имеется еще одна оболочка - покрывалъце. Внутри лежит гимениальный слой, состоящий из развивающихся сумок и парафиз. Этот слой формируется на узком внутреннем слое стенки перитеция - гипотеции. Вблизи отверстия имеются особые нитевидные гифы - перифизы, выполняющие защитную функцию.
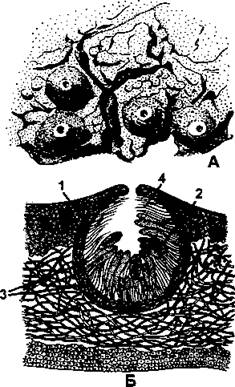
Рис. 398. Перитеции:
А - внешний вид перитециев; Б - поперечный разрез через слоевище с погруженным в него перитецием (1 - эксципул; 2 - гимениальный слой, образованный сумками со спорами и парафизами; 3 - гипотеций; 4 - перифизы) (по Н. С. Голубковой)
Форма сумок может быть самой разнообразной (рис. 399), количество созревающих в них спор у разных видов также неодинаково - от одной крупной до 200.
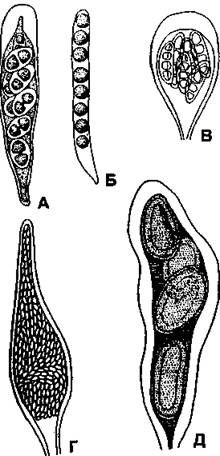
Рис. 399. Форма сумок лишайников:
А - удлиненно-булавовидная (Buellia);
Б - цилиндрическая (Sphaerophorus);
В - булавовидная (Arthonia);
Г - бутылковидная (Thelocarpon);
Д - мешковидная (Pertusaria)
(по Н. С. Голубковой)
У большинства видов стенка сумки имеет одну оболочку (унитуникатные - лат. unus, uni - один, tunica - наружный слой, покров), но у некоторых форм (прежде всего у асколокулярных лишайников) оболочек может быть две (битуникатные - лат. bi - дву(х), tunica - наружный слой, покров) (рис. 400).
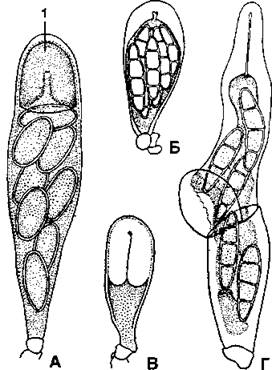
Рис. 400. Сумки лишайников:
А - унитуникатные сумки с апикальным аппаратом (1) на вершине, с помощью которого споры высвобождаются из сумок;
Б, В - зрелая и молодая битуникатные сумки;
Г - выбрасывание спор из битуникатной сумки (по Н. С. Голубковой)
Споры окружены двуслойной оболочкой, состоящей из наружного слоя (экзоспория) и внутреннего (эндоспория). Каждая спора содержит ядро и небольшое количество цитоплазмы. Форма и размеры спор разнообразны (рис. 401). Выход спор может быть пассивным в результате разрушения стенки сумки, но чаще они выбрасываются из сумки за счет увеличения внутри сумки осмотического давления. Общее количество спор, образуемых лишайником, очень велико. Например, только один апотеций лишайника солорины продуцирует около 124 000 спор. Выход спор из сумок зависит от погодных условий и прежде всего влажности - увлажнение стимулирует освобождение спор (споруляцию).
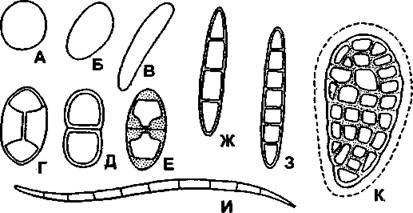
Рис. 401. Споры лишайников:
А - шарообразные;
Б - эллиптические;
В - удлиненные;
Г - биполярные;
Д - двухклеточные;
Е - двухклеточные с сильно и неравномерно утолщенными оболочками;
Ж - четырехклеточные вытянутые;
З - восьмиклеточные вытянутые;
И - игловидные многоклеточные;
К - муральные (по Н. С. Голубковой)
Большинство спор погибает, и только немногие из них попадают в благоприятные условия и прорастают.
Проросшие из споры гифы выживают только, если встретят поблизости подходящую водоросль. Тогда клетки водоросли оплетаются гифами гриба и постепенно формируется таллом (см. рис. 377).