В эту гетерогенную группу объединены все грибы с членистыми гифами, но с неизвестным до настоящего времени половым процессом. Экспериментально установлено, что рекомбинация генетического материала у них осуществляется в ходе гетероядерности и парасексуалъного процесса (Дж. Понтекорво). Гетороядерность возникает в тех случаях, когда разнородные ядра переходят из одной гифы в другую. Если они после этого сливаются, возникает диплоидное ядро с разными аллелями. Дальнейшие митотические деления этого ядра обогащают генетическое разнообразие организма, причем измененные ядра могут опять стать гаплоидными в случае потери хромосом. Таким образом, рекомбинация аллелей у несовершенных грибов происходит в процессе митоза, а не мейоза.
Бесполое размножение осуществляется конидиями и систематическая характеристика (совершенно условная) базируется на форме конидий и расположении конидиеносцев. Представители этой группы имеют неодинаковое происхождение, поэтому их еще называют формальной группой. Несовершенные грибы чрезвычайно широко распространены в природе, среди них есть сапрофиты и паразиты, некоторые из них приводят к гибели высших растений. Всего насчитывается более 30 000 видов несовершенных грибов. Согласно наиболее распространенной систематике итальянского миколога П. А. Саккардо, различают три группы несовершенных грибов: Гифомикоты, Мелянкониевые и Сферопсидные.
Гифомикоты являются самой крупной группой несовершенных грибов. Конидиеносцы у них могут быть одиночными, или они собраны в группы - пучки (коремии) или сплетения (спородохии). Различные виды гифомикот могут быть наземными или водными, а по типу питания сапрофитными и паразитами. Среди сапрофитов имеются виды, которые продуцируют антибиотики, и поэтому используются человеком в биотехнологическом производстве. Паразиты растений (фитопаразиты) вызывают заболевания высших растений, которые могут существенно снижать урожаи (например, гриб ботритис, вызывающий серую гниль различных частей растений). Интересную группу составляют хищные гифомикоты, которые способны улавливать различных беспозвоночных (простейших, коловраток, нематод и даже мелких насекомых) и в дальнейшем питаться ими. Для поимки своих жертв хищные грибы формируют ловушки из особых гиф (рис. 370).

Рис. 370. Типы ловушек хищных грибов:
А - клейкие трехмерные сети;
Б, Е - клейкие головки:
В - сжимающиеся кольца:
Г - клейкие выросты гиф;
Д - несжимающиеся кольца (по И. И. Сидоровой)
Меланкониевые формируют плотный слой переплетенных конидий на более или менее плоском ложе (рис. 371). Большинство видов (а их всего около 1000) являются сапрофитами, некоторые ведут паразитический образ жизни, поражая надземные части растений. Они вызывают заболевания пятнистость и антракноз.

Рис. 371. Строение ложа меланкониевых грибов (по Л. Н. Левкиной)
Сферопсидные (пикнидиальные) образуют конидиеносцы внутри особых споровместилищ - пикнид, которые имеют шаровидную, овальную или грушевидную форму с отверстием на вершине (рис. 372). Споры (их также называют стилоспорами, или пикноспорами) выходят из пикнид через имеющиеся на поверхности отверстия. Сферопсидные грибы широко распространены, кроме районов Крайнего Севера и Антарктиды. Часть из них сапрофиты, другие - паразиты высших и низших растений, грибов и лишайников. Многие виды вызывают заболевания культурных растений, например фому свеклы, белую гниль винограда, аскохитоз гороха, сухую гниль кукурузы и др. Всего известно около 6000 видов сферопсидных грибов.
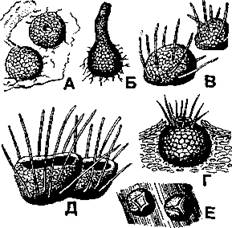
Рис. 372. Пикниды родов:
А - фома (Phoma);
Б - пленодом (Plenodomus);
В - хетомелла (Chaetomella);
Г - войновиция (Wojnowicia);
Д - динемаспориум (Dinemasporium);
Е - дисцелла (Discella) (по Г.Д. Успенскому)