ГОМОЛОГИЧЕСКИЕ РЯДЫ НАСЛЕДСТВЕННОЙ ИЗМЕНЧИВОСТИ. ВНЕЯДЕРНАЯ НАСЛЕДСТВЕННОСТЬ
Термины и понятия: «один ген — один признак»; признаки качественные (моногенные) и количественные (полигенные); «один ген — одна полипептидная цепь»; закон гомологических рядов наследственной изменчивости; цитоплазматическая наследственность.
Классическая концепция: «один ген —один признак». В период развития классической генетики (первая половина XX ст.), которая ещё не основывалась на данных молекулярной биологии, основным генетическим правилом считали: «один ген — один признак». Это значит, что какое-либо наследственное изменение признака или появление нового признака связано исключительно с изменением существующего гена или появлением нового.
По мере проводимых исследований выяснилось, что моногенных (от греч. монос — один и ген) фенотипических признаков, изменчивость которых обусловлена одним геном, в природе не так уж много. Большинство признаков полигенны (от греч. полис — много и ген) и определяются двумя и более независимыми генами, каждый из которых представлен двумя или несколькими аллелями. Моногенные признаки обладают качественным проявлением (цвет глаз или волос, курчавость, ранняя плешивость людей, белая и красная окраска цветов, признаки гороха, взятые Менделем для исследования). Именно поэтому их часто называют качественными. Изменчивость полигенных признаков оценивается количественно. Если это размерные характеристики, то соответственно используются единицы измерения, если это непрерывно меняющиеся физиологические показатели устойчивость к болезням, продуктивность, жирность молока у коровы и т. д.), то они оцениваются в особых единицах. Не случайно непрерывно изменяющиеся признаки принято называть количественными.
Давно известно, что у большинства организмов, обитающих в дикой природе, очень редко проявляется изменчивость по качественным признакам. Крайне редко в природе можно увидеть белую ворону или охотнику добыть чёрно-бурую лисицу. Обычно все организмы в природной среде обитания представлены совершенно определённым фенотипом — диким типом, свойственным подавляющему большинству особей этого вида. Особи дикого типа тоже отличаются друг от друга внешними признаками, но только количественными. Если речь идёт о вороне, то это значит, что одна особь крупнее, другая — мельче, у одной клюв длиннее, у другой — уже, у третьей он — и массивнее и так далее. Эти отличия особей друг от друга во многом могут формироваться не столько генотипами генов, определяющими развитие этого признаки, сколько действием различных факторов среды обитания. Дикий тип формируется доминантными аллелями, которые находятся в гомозиготном или гетерозиготном состоянии. Выщепление рецессивных генов в гомозиготу и приводит к появлению белых ворон среди чёрно-серых (рис. 63).

Рис. 63. Редкий случай вороны с мутантным фенотипом белой окраски.
В отличие от естественной среды обитания, при разведении животных в искусственных условиях в их популяциях накапливается большое количество уклоняющихся от дикого типа фенотипов, обусловленных гомозиготными сочетаниями рецессивных генов. Достаточно сравнить разнообразные окраски полудиких городских голубей, с серо-стальной с переливами окраской их диких предков (рис. 64). В скальных популяциях голубей не встретишь ни белых, ни пёстрых, ни рыжих птиц. Почти полное отсутствие в природе мутантных особей связано не только с тем, что, например, белой вороне трудно подыскать пару для размножения или её быстрее выследит хищник на дереве, но ещё и с тем, что любая мутация обладает плейотропным действием. Причём эффект от действия этого гена может внешне никак не проявляться, хотя внутренние разлады, например, нарушения обмена веществ, как раз и приводят к тому, что в морозные зимы при дефиците корма, у белой вороны гораздо меньше шансов выжить, чем у особей дикого типа.

Рис. 64. Голубь «дикого типа» (а) и разные мутантные фенотипы (б—г).
Современная концепция: «один ген — одна полипептидная цепь» Когда стало ясно, что основная функция генов заключается в том, чтобы служить матрицей для синтеза определённого типа РНК, причём часть этих РНК, в свою очередь, служит матрицей для синтеза полипептидной цепи, было сформулировано ещё одно генетическое правило: «один ген — одна полипептидная цепь». Ведь стало очевидным, что все гены, так или иначе определяют, какие белки и в каком количестве будут синтезироваться в клетке. Последнее больше относится к регуляторным генам, функция которых состоит в регуляции активности структурных генов. Регуляторные гены не только определяют скорость синтеза РНК на тех или иных последовательностях ДНК, но даже способны переключать гены — останавливать или включать транскрипцию.

Рис. 65. Изменчивость раковины лунки речной вызвана модификацией генов окраски: пятнистый (а)и полосатый (б) фенотипы.
Следует сказать, что если на структурные гены среда не оказывает непосредственного влияния, то на генную регуляцию вполне можно воздействовать с помощью определённых факторов среды. В результате изменяется экспрессия (от англ. expression — выражение) — активность тех или иных генов, что может иметь и явные фенотипические эффекты. Один из самых интересных примеров — изменчивость пёстрой окраски раковины лунки речной (рис. 65). Этот небольшой моллюск обитает почти во всех реках Украины. Оказывается, поперечно полосатую окраску на пятнистую и обратно можно менять в ряду поколений, варьируя температуру, pH, содержание солей в воде, в которой развивается моллюск, или просто на какое-то время подавляя его рост. Такие модификации возможны только у особей, имеющих одновременно гены, отвечающие за тот или иной тип окраски. В определённых условиях действие гена пятнистой окраски доминирует над поперечнополосатой, в иных условиях достигается обратный результат.
В конце концов, стало ясно, что генетическая уникальность каждого организма или вида реализуется в особенностях набора белков, а также в том, как регулируется активность генов. Следует учесть, что белки — не только строительный материал клетки, но и ферменты, катализирующие все химические реакции в организме. Очевидно, что как нарушение первичной структуры полипептидной цепи, вызванное мутациями конкретного гена, так и уменьшение его активности приведут к дефициту функции белка в клетке. Это может стать причиной самых различных отклонений в строении клетки или нарушений обмена веществ. В последнем случае не будут синтезироваться нужные вещества (вспомните генетические механизмы альбинизма) и накапливаться ненужные (вспомните, что вызывает фенилкетонурию).
Проиллюстрировать, каким образом мутации в разных генах приводят к сбоям в метаболизме и фенотипическим эффектам, можно на примере генов, определяющих разный цвет глаз дрозофилы. Выделяют серию аллельных генов, каждый из которых имеет своё фенотипическое проявление. Кроме двух крайних фенотипов: красные глаза (доминантный ген) дикого типа и полное отсутствие пигмента — белые глаза (рецессивный ген), был обнаружен ряд фенотипов, определяемых серией аллелей: алые глаза, розовые глаза, абрикосовые глаза, жёлтые глаза. Эти гены последовательно доминируют один над другим в зависимости от того, насколько цвет глаз, который они определяют, приближён к дикому типу.
Оказалось, что появление этой серии аллелей вызвано мутациями структурных генов, которые кодируют разные ферменты, катализирующие последовательный ряд химических реакций синтеза меланина. Если в гомозиготном сочетании окажутся мутантные гены фермента, осуществляющего самую первую реакцию на пути синтеза меланина, то это приведёт к полной остановке образования не только меланина, но и меланиноподобных соединений, а потому глаза у дрозофилы будут, как у настоящего альбиноса, белыми. Если в гомозиготном состоянии будут иметь место мутации, затрагивающие гены, которые кодируют ферменты второй, третьей, четвёртой реакций на пути синтеза меланина, то на соответствующих его стадиях будут синтезироваться производные вещества, окрашивающие глаза в тот или иной цвет. Причём по мере превращений химические соединения по своим формулам всё более будут напоминать меланин, а цвет глаз всё более насыщаться красным. И, наконец, у особей дикого типа с красными глазами будет либо гомозиготное сочетание доминантных аллелей, определяющих цвет глаз, либо любое гетерозиготное сочетание с участием доминантного аллеля.
Похожая ситуация и с цветом глаз у человека. Мутации структурных генов, которые служат матрицами для синтеза ферментов, катализирующих синтез меланина и преобразование других пигментов, в конечном счёте определяют разную степень насыщения радужной оболочки меланином в следующей последовательности: голубые, серые, зелёные, янтарные, болотные, карие глаза. При этом гены с фенотипическим эффектом большего насыщения меланина являются доминантными по отношению к менее насыщенным.
Остальные фенотипические действия генов также могут быть интерпретированы как проявление различных мутаций белковых локусов, как правило, в гомозиготных сочетаниях генов, которые приводят к тому, что некоторые реакции в клетках перестают происходить или совершаются недостаточно быстро. Если это ключевые реакции (например, обеспечивающие репликацию, транскрипцию, трансляцию, энергетический обмен), то это ведёт к летальному исходу на самых ранних стадиях развития, если это реакции вторичного метаболизма (к этой категории относится синтез всевозможных пигментов), то это вызовет разнообразные фенотипические проявления (в данном случае изменение цвета).
Гомологические ряды мутационной изменчивости. Таким образом, причиной очень многих мутаций с внешним фенотипическим проявлением являются сбои в метаболизме, вызванные мутациями генов, кодирующих ферменты. Так как метаболические циклы и пути даже у отдалённых видов очень похожи (вспомните: у всех организмов по одному принципу происходит биосинтез ДНК, РНК, белков, а у всех растений, грибов и животных обязательно есть гликолиз и цикл Кребса), то получается, что у разных видов неизбежно возникают одинаковые нарушения метаболизма, вызванные мутациями одних и тех же структурных генов. Если это близкие, внешне очень похожие виды с одинаковыми особенностями онтогенеза, то и фенотипический эффект от этих мутаций будет проявляться похоже. Возникнет, как принято говорить, параллельная изменчивость (рис. 66).

Рис. 66. Параллельная изменчивость остистости колосьев: 1 —4 — мягкая пшеница; 5—8 — твёрдая пшеница; 9—12 — ячмень.
Проявления одинакового спектра мутаций у близких видов организмов были названы гомологичными рядами наследственной изменчивости и сформулированы в 1920 г. выдающимся генетиком, академиком Национальной академии наук Украины Николаем Ивановичем Вавиловым (1887—1943) следующим образом: целые семейства растений, в общем, характеризуются определённым циклом изменчивости, проходящей через все роды и виды. В настоящее время это положение часто называют законом гомологических рядов наследственной изменчивости.
Сейчас закон трактуют шире и распространяют не только на растения, но и на животных и даже бактерии. Типичным примером параллельной изменчивости является мутация альбинизма, характерная не только для млекопитающих, птиц, рыб, но даже растений. Закон гомологических рядов имеет важное теоретическое и практическое значение, поскольку позволяет предсказать: если у одного из видов обнаружена определённая мутация, то она должна обязательно проявиться и у других видов этого рода.

Рис. 67. Н. И. Вавилов.
Внеядерная, или цитоплазматическая, наследственность. В ряде случаев при изучении наследования моногенных признаков был установлен ряд необычных отклонений от менделевского наследования, связанных с полом. Исследования показали, что причина этого явления в том, что мутации происходят не в генах хромосом, а в генах, находящихся в самовоспроизводящихся органеллах — митохондриях и пластидах (вспомните: эти органеллы точно так же, как и клетки, размножаются делением пополам). Действительно, у пластид и митохондрий имеется свой генетический аппарат — свёрнутая в кольцо двухцепочная молекула ДНК, напоминающая бактериальную хромосому.
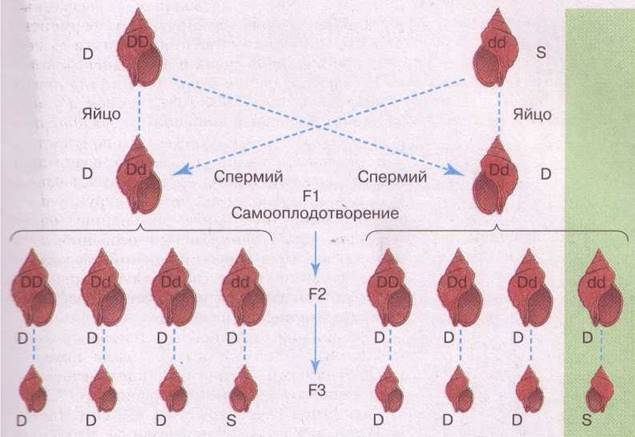
Рис. 68. Схема наследования направления завитка прудовика большого. Фенотипы: D — правосторонний и S — левосторонний; аллели: D — доминантный, определяющий правосторонний завиток, и d — рецессивный, определяющий левосторонний завиток.
Поскольку пластиды и митохондрии передаются потомству только через цитоплазму яйцеклетки (у сперматозоидов практически нет цитоплазмы), то соответственно все генетические свойства, закодированные в генетическом материале этих органелл, передаются по материнской линии. Этот способ внеядерной передачи генетического материала получил название цитоплазматической наследственности, которая реализуется только по материнской линии.
Одним из классических примеров цитоплазматического наследования является передача особенностей формы раковины у моллюска прудовика большого (вспомните: вы его изучали на уроках биологии в 8 классе). У одних рас моллюсков раковина завивается вправо — признак обусловлен доминантным геном, а у других влево — признак обусловлен рецессивным геном. Как выяснилось, в первом поколении гибридных скрещиваний направление завитка у потомства задаётся не генотипами улиток, а материнским организмом. Это связано с тем, что второе деление митоза при дроблении, определяющее направление завитка, обусловлено организацией цитоплазмы клетки. В результате в первом поколении независимо от того, какой ген — рецессивный, а какой — доминантный, потомство будет иметь такой же завиток, что и материнской организм, и только во втором поколении произойдёт расщепление в строгом соответствии со вторым законом Менделя.
Теперь, вооружившись этими знаниями, на вопрос: «На кого ты больше похож - на маму или папу?», можете смело отвечать: «На маму. У меня от неё митохондриальная ДНК».
Кроме того, в цитоплазме клеток есть особые генетические факторы плазмиды — представляющие собой небольшие линейные или замкнутые в кольцо молекулы ДНК. Лучше всего плазмиды изучены у бактерий. Именно в бактериальных плазмидах содержатся гены, обеспечивающие бактериям устойчивость к антибиотикам или определяющие их пол.
Одним из главных принципов современной генетики является положение: «один ген — одна полипептидная цепь». Согласно ему мутации в генах приводят к нарушениям структуры белков и ферментов, следствием чего становятся нарушения метаболизма, что сопровождается фенотипическим проявлением.
Близкие виды организмов имеют сходный набор мутаций структурных генов, проявляющихся фенотипически в виде гомологических рядов наследственной изменчивости.
Не вся генетическая информация находится в хромосомах, часть её содержится в генетических аппаратах митохондрий, пластид или в виде небольших молекул ДНК в цитоплазме клетки. Этот генетический материал передаётся по материнской линии. Такой способ передачи информации получил название цитоплазматической наследственности.
Проверьте себя
1. Почему в природе так редко встречаются белые вороны? 2, В чём суть концепции: «один ген - одна полипептидная цепь»? 3. Каким образом мутации генов, кодирующих ферменты, приводят к фенотипическим эффектам? 4. Что называют параллельной изменчивостью? 5. Что собой представляют факторы цитоплазматической наследственности?
Как вы считаете?
1 Почему домашние животные, попавшие в дикую природу и образующие там популяции, с каждым поколением всё больше возвращаются к дикому типу? Возможен ли в таком случае полный возврат к дикому типу?
2. Почему гены, обусловливающие цвет глаз с большим насыщением меланина, доминируют над генами, обусловливающими цвет глаз с меньшим его количеством?