Обнаружение и идентификация опухолевых антигенов
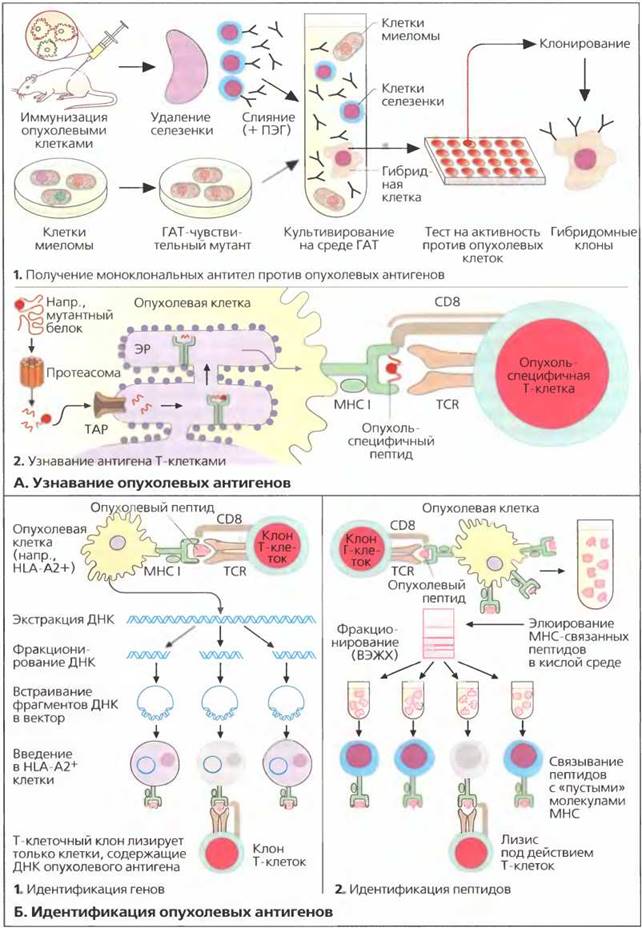
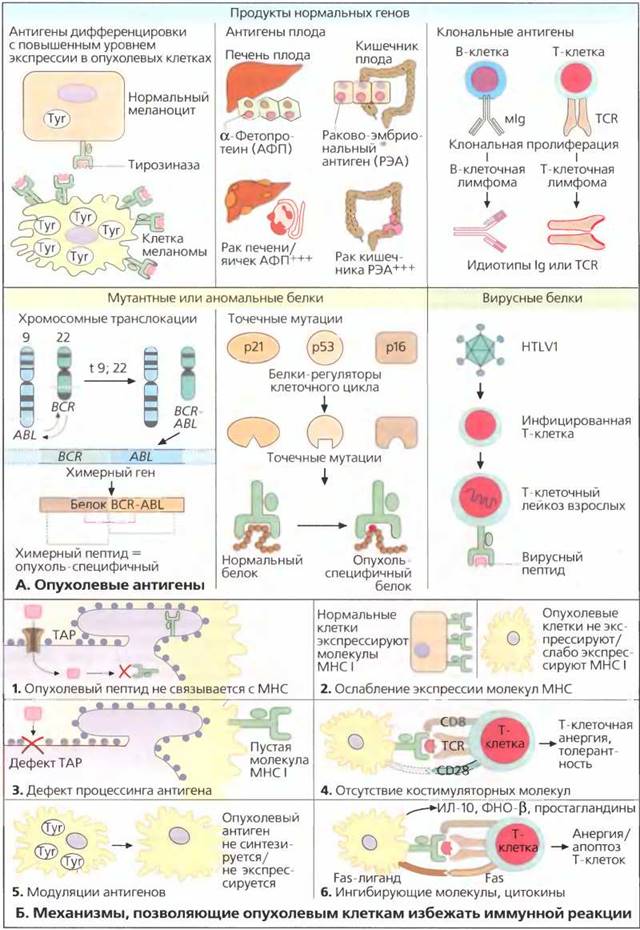
А. Опухолевые антигены
В идеальном варианте для того чтобы быть узнанным иммунной системой в качестве чужеродного антигена, опухолевый антиген должен экспрессироваться только опухолевыми клетками, но не нормальными клетками организма. В реальной ситуации большинство опухолевых антигенов очень активно экспрессируется опухолевыми клетками, но в меньших количествах экспрессируется и нормальными клетками. Например, фермент тирозиназа экспрессируется всеми нормальными меланоцитами, но в клетках меланомы ее концентрация гораздо выше. Специфичные к тирозиназе Т-клетки, распознающие и уничтожающие клетки меланомы, обнаруживаются в крови некоторых пациентов с меланомой. Эти Т-клетки на самом деле ответственны за депигментацию кожи (витилиго) у некоторых пациентов. Наличие витилиго часто коррелирует с реакцией на терапию.
Онкофетальные антигены (например, альфа-фетопротеин или раково-эмбриональный антиген (РЭА)) присутствуют в опухолевых клетках печени и гонад, а также различных аденокарцином. Эти антигены активно экспрессируются в процессе развития плода, но в норме слабо экспрессируются во взрослом организме. Онкофетальные антигены циркулируют в крови больных раком в виде растворимых белков и поэтому могут использоваться как индикаторы прогрессии опухолей (опухолевые маркеры).
При В- и Т-клеточных лейкозах и лимфомах клон- специфичные детерминанты (идиотипы) иммуноглобулинов и Т-клеточных рецепторов представляют собой индивидуальные и специфичные опухолевые антигены. Следовательно, они не должны участвовать в реакциях в других тканях.
С другой стороны, мутантные белки, возникающие в результате хромосомных транслокаций или точечных мутаций, представляют собой совершенно новые антигены (неоантигены). Например, при хроническом миелолейкозе имеет место транслокация t(9:22), приводящая к образованию химерного белка BCR-ABL. Пептиды с аминокислотными последовательностями, соответствующими обоим генам BCL и ABL, могут действовать как истинные опухолевые антигены. Точечные мутации, которые часто возникают в последовательности гена- супрессора опухоли р53 и в генах других белков-регуляторов клеточного цикла [р16 и р21), также приводят к образованию новых опухоль-специфичных пептидов, не существующих в нормальных клетках. Однако для того чтобы быть узнанными иммунной системой, эти пептиды должны соответствовать молекулам HLA в опухолевых клетках. Следовательно, могут существовать опухоли, в которых присутствуют такие специфические пептиды, но они не презентируются иммунной системе индивидуальными молекулами МНС. Истинные вирусные белки, образуемые вирусом HTLV1 при Т-клеточ- ном лейкозе и вирусом Эпштейна-Барр при некоторых злокачественных лимфомах, ответственны за злокачественную трансформацию и синтезируются опухолевыми клетками. Очевидно, что такие вирусные белки могут успешно узнаваться иммунной системой. В отличие от белков с единичными мутациями, такие вирусные белки обычно содержат много потенциально иммуногенных пептидов, и поэтому существует большая вероятность, что некоторые из них будут связываться с молекулами МНС организма хозяина.
Б. Механизмы, позволяющие опухолевым клеткам избежать иммунной реакции
Существует ряд причин, объясняющих отсутствие эффективного иммунного ответа на опухолевые клетки. Каждый аутологичный белок разлагается внутри цитоплазмы с образованием пептидов, состоящих из 9-12 аминокислотных остатков. Эти пептиды препровождаются в эндоплазматический ретикулум (ЭР) специальными транспортными белками, называемыми транспортерами, связанными с процессингом антигена (ТАР). В ЭР пептиды связываются молекулами МНС класса I и презентируются CD8+ Т-лимфоцитам на поверхности клетки (см. с. 66).
В некоторых опухолях может не быть пептидов, соответствующих участкам связывания на молекулах МНС больного (рис. Б1). В опухолевых клетках с нарушением механизма процессинга антигена (например, с дефицитом ТАР) опухолевые пептиды не транспортируются в ЭР (рис. БЗ) и не происходит их презентации на поверхности клетки. Во многих случаях происходит потеря молекул МНС класса I на поверхности опухолевых клеток, связанная со снижением экспрессии генов МНС. В результате опухолевые антигены не распознаются цитотоксическими Т-клетками (рис Б2).
Опухолевые клетки не являются профессиональными антигенпрезентирующими клетками. У них нет костимулирующих молекул CD80 и CD86, необходимых для активации Т-клеток. Без костимуляции презентация пептида посредством комплекса MHC/TCR приводит к анергии и толерантности Т-клеток (рис. Б4) Некоторые опухолевые клетки прекращают синтезировать опухолевые антигены, чтобы избежать иммунной реакции (рис. Б5). Кроме того, опухоль может производить иммуносупрессоры типа ИЛ-10 и трансформирующего фактора роста β (ТФР-(3) (рис. ББ), а в некоторых случаях — и МНС l-подобные молекулы, которые взаимодействуют с ингибирующими лигандами на Т-клетках. Недавно было показано, что регуляторные Т-клетки (CD4+, CD25+) подавляют иммунный ответ у пациентов с солидными опухолями
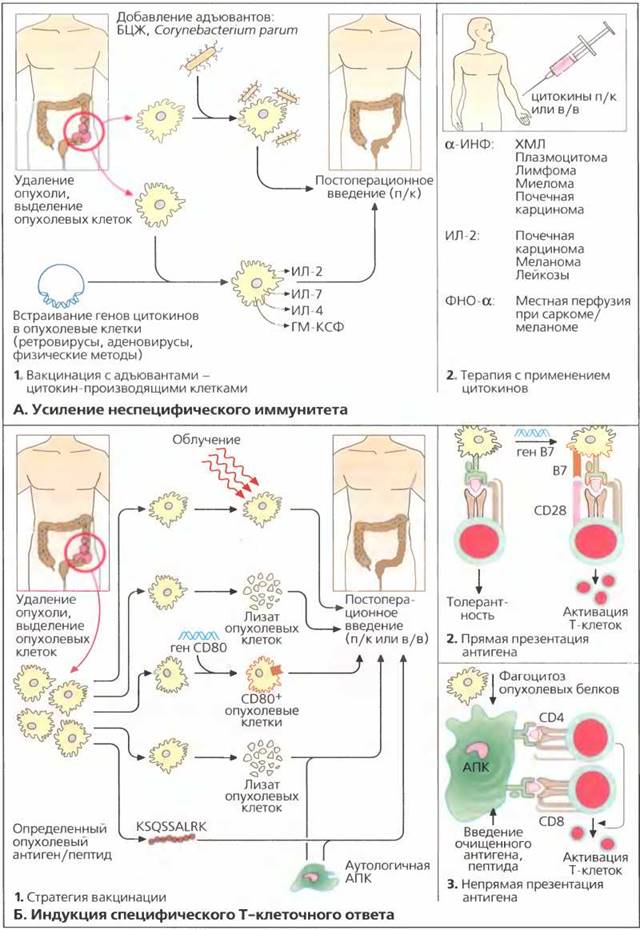
Преодоление иммунологического надзора опухолевыми антигенами
А. Усиление неспецифического иммунитета
В конце XIX в. американский хирург Уильям Колей начал вводить раковым больным лизаты опухолевых клеток, которые были заражены бактериями из-за неразработанности методик стерильной работы. Удивительно, но у некоторых пациентов происходила регрессия опухоли. Позднее, уже в XX в. опыт с противоопухолевой вакцинацией был вновь повторен, однако использование только немодифицированных опухолевых клеток не приводило к возникновению ощутимого иммунного ответа. Вскоре в вакцину стали добавлять адъюванты. Сначала использовали ослабленные штаммы микобактерий: бациллу Кальметта-Герена (БЦЖ) и Corynebacterium parvum. В более поздних экспериментах для этой цели пытались использовать вирус ньюкастлской болезни. До сих пор ведутся некоторые клинические испытания, основанные на данной стратегии лечения. Стремительного успеха от этих исследований ожидать трудно, но некоторые положительные данные указывают на возможность применения усовершенствованных методов вакцинации в будущем.
В последние три-четыре года были проведены работы по генетической модификации опухолевых клеток с целью достичь селективной индукции Т-клеточного иммунного ответа. Ставилась задача стимулировать локальный рост Т-клеток или дендритных клеток путем секреции цитокинов (ИЛ-2, ИЛ-4, ИЛ- 7 и ГМ-КСФ) в надежде активировать специфичные к опухоли Т-клетки.
В терапевтических целях можно также использовать системное введение цитокинов. Однако на практике при отдельных видах опухолей используются только а-интерферон и ИЛ-2 (рис. А2). Кроме иммуностимулирующей активности, а-ИНФ обладает и прямым антипролиферативным действием, что может объяснять наблюдаемый терапевтический эффект. Лучше остальных видов опухолей на иммунотерапию реагируют почечные карциномы и злокачественные меланомы, однако причина этого явления не ясна При лечении пациентов с саркомами и меланомами при перфузии конечностей для усиления воздействия на опухолевые клетки используется также фактор некроза опухолей ФНО-а в сочетании с противоопухолевыми препаратами.
Б. Индукция специфического Т-клеточного ответа
У многих пациентов хирургическое вмешательство в момент постановки диагноза может успешно уничтожить первичную опухоль. Однако у некоторых из этих пациентов через какое-то время будут образовываться отдаленные метастазы или произойдет повторное возникновение опухоли. Вот почему пациентам с определенными факторами риска (тип опухоли, степень злокачественности, глубина проникновения опухоли, вовлечение лимфатических узлов и др.) показано проведение адъювантной терапии. Тем не менее в большинстве случаев пока не удается индуцировать специфический Т-клеточный ответ на опухоль путем введения облученных аутологичных опухолевых клеток или лизатов опухолевых клеток (рис. Б1). Даже в идеальной ситуации, когда опухолевые клетки осуществляют презентацию опухолевого антигена Т-клеткам, сильный иммунный ответ не достигается из-за отсутствия у опухолевых клеток костимуляторных молекул типа В7 (CD80/CD86) (см. с. 46 и 163). Проведен целый ряд испытаний генетически модифицированных опухолевых клеток, экспрессировавших В7 для усиления иммуногенности (рис. Б2). Еще одним способом активизации иммунного ответа на слабоиммуногенные опухолевые клетки является презентация опухолевого антигена специализированными антиген- презентирующими клетками (АПК) (рис. БЗ). Специализированные АПК экспрессируют все необходимые костимуляторные молекулы, необходимые для индукции эффективного Т-клеточного ответа Таким образом, вместе с лизатами опухолевых клеток, очищенными опухолевыми антигенами или специфическими опухолевыми пептидами можно ввести дендритные клетки, образованные ex vivo. На сегодняшний день в этой области исследований получен ряд обнадеживающих результатов.
Особой формой иммунотерапии является капельное введение вакцины БЦЖ после локальной резекции карциномы мочевого пузыря. БЦЖ в значительном числе случаев предотвращает повторное возникновение опухоли. Введение бациллы может запускать воспалительный процесс, в результате которого активированные антигенпрезентирующие клетки способны захватывать антигены оставшихся опухолевых клеток и эффективно осуществлять их презентацию
Иммунотерапевтические стратегии (I)
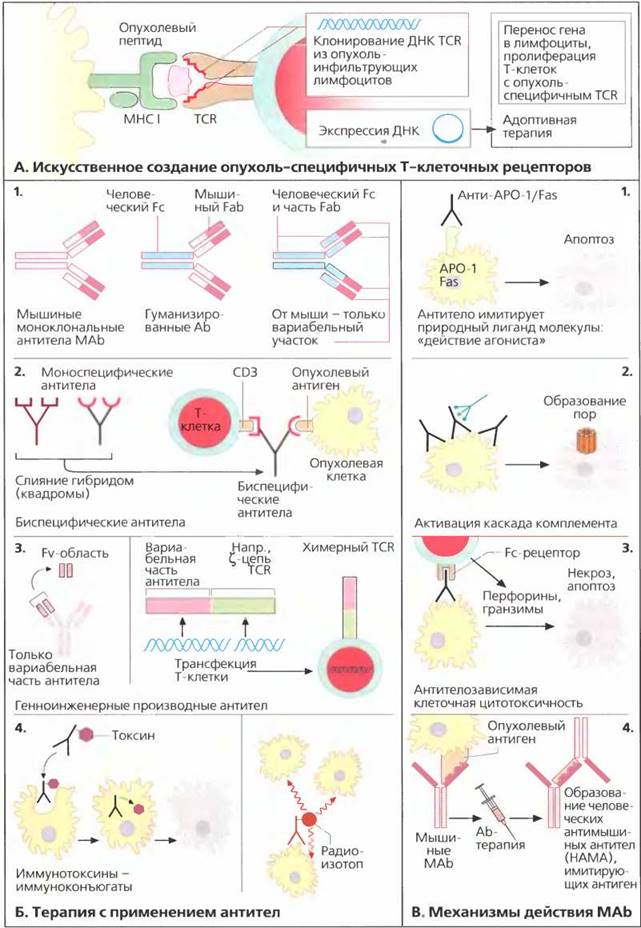
А. Генноинженерные опухоль-специфичные Т-клеточные рецепторы
Многие опухоли интенсивно инфильтруются лимфоцитами. Были предприняты попытки выделять инфильтрующие опухоль лимфоциты TIL из удаленных опухолей, особенно из меланом и почечных карцином, и вновь переливать их пациенту. Однако результаты оказались скорее обескураживающими. Теперь возможно определить последовательность ДНК вариабельных доменов а- и β-цепей Т-клеточных рецепторов TIL. С помощью вирусных векторов можно получить генетически измененные Т-лимфоциты, специфическим образом реагирующие с опухолевыми клетками Затем следует осуществить адоптивную терапию, вводя пациенту эти размноженные ex vivo клетки. В настоящее время наибольшую проблему представляет этап переноса генов в нормальные Т-клетки, однако постоянно ведется интенсивный поиск новых методов. Также необходимо обеспечить снижение экспрессии эндогенных Т-клеточных рецепторов.
Б. Терапия с применением антител
Моноклональные антитела (МАb) используются для лечения рака уже на протяжении более 20 лет. Большинство антител получают в мышах. Лечение мышиными антителами приводит к образованию человеческих анти-мышиных антител (НАМА), что снижает эффективность мышиных МАb. В связи с этим была разработана технология получения гуманизированных МАb (рис. Б1). В гуманизированных антителах большая часть молекулы имеет человеческую природу, и только Fab- фрагмент или вариабельный участок Fab-фрагмента взяты от мыши. Гуманизированные моноклональные антитела имеют гораздо большее время полувыведения из крови пациента и гораздо эффективнее активируют иммунную систему по сравнению с мышиными антителами. В последние несколько лет прекрасные результаты были получены при применении двух типов гуманизированных моноклональных антител: анти-СD20 антитела ритуксимаб (препарат ритуксан) обладают активностью против В-клеточных лимфом, а антитела трастузумаб (препарат герсептин), направленные против рецептора человеческого эпидермального фактора роста 2 (HER2), обладают активностью при раке молочной железы. Получен ряд обнадеживающих результатов и с другими антителами. Еще одним новым подходом является использование биспецифических антител (рис. Б2). Биспецифические антитела образуются путем слияния двух гибридом, каждая из которых секретирует специфические моноклональные антитела. Такие гибридомы одновременно связывают эпитопы опухолевых клеток и Т-клетки (CD3). Таким образом, Т-клетки вступают в контакт с опухолевыми клетками и активируются при помощи CD3. Новшеством в данной стратегии стало использование синтетических антител, состоящих из одной цепи (рис. Б3). Эти полученные генноинженерным способом антитела состоят только из легкой и тяжелой цепи вариабельной области (Fv) МАb. Единичные цепи от двух различных антител могут быть скреплены при помощи связывающего фрагмента (спейсера]. В альтернативном варианте состоящие из одной цепи антитела, узнающие опухолевый эпитоп, могут быть связаны с дзета-цепью, играющей важную роль при передаче сигнала TCR. Можно осуществить трансфекцию Т-клеток такими конструкциями, в результате чего они смогут узнавать опухолевый антиген таким же образом, как это делают антитела, и передавать сигнал внутрь клетки при помощи дзета-цепи TCR. Антитела, направленные против опухолевых антигенов, можно связать с иммунотоксинами или радиоизотопами (рис. Б4). Токсический компонент иммунотоксина, блокирующий синтез РНК, высвобождается в результате интернализации антител. В случае меченных радиоактивными изотопами конъюгатов радиоактивное вещество (обычно радиоактивный иттрий или иод) вызывает лизис клетки-мишени, но при этом также страдают расположенные рядом клетки (эффект «невинного свидетеля»)
В. Механизмы терапевтического действия моноклональных антител
Моноклональные антитела могут имитировать или блокировать естественные лиганды рецепторов. Например, анти-С095 антитела активируют рецептор Fas/APO-1 (CD95), запуская апоптоз (рис. В1). С другой стороны, антитела могут активировать комплемент и индуцировать образование пор в клеточной мембране (рис. В21 При антителозависимой клеточной цитотоксичности клетки-киллеры, несущие Fc-рецепторы, узнают Fc-фрагмент связанных с клеткой антител, что приводит к высвобождению цитоплазматических гранул, содержащих цитотоксические перфорины и гран-зимы (рис. В3, см. также с. 49).
Как уже говорилось, мышиные антитела могут индуцировать образование НАМА. Некоторые из этих анти-мышиных антител могут быть направлены против специфических идиотипических связывающих областей мышиных антител (рис. В4). В таком случае анти- идиотипические антитела имитируют опухолевые антигены и могут служить опухолевыми суррогатами в вакцинах. С другой стороны, они также могут приводить к образованию анти-анти-идиотипических моноклональных антител, которые узнают опухолевый антиген, подобно мышиным МАb. В результате реализации этого каскада действие мышиных моноклональных антител усиливается
Иммунотерапевтические стратегии (II)