Слизевики, или миксомицеты (греч. шуха - слизь, mykes, myketos — гриб), представляют собой группу своеобразных организмов, сочетающих в себе признаки животных и грибов. Часть своей жизни они проводят в виде отдельных амебоподобных или жгутиковых клеток, часть - в виде многоядерного плазмодия (вегетативного тела), кроме того, слизевики образуют плодовые тела самой разнообразной формы.
Морфологически вегетативное тело представляет собой более или менее крупную цитоплазматическую массу, содержащую большое количество ядер. Внешняя жесткая оболочка отсутствует, поэтому перетекание цитоплазмы из одного участка плазмодия в другой обеспечивает медленное поступательное движение. Этим же объясняется непостоянство формы плазмодия. Для плазмодия характерно высокое содержание белков - до 30% всей массы, из запасных углеводов присутствует гликоген, однако свойственный растениям крахмал отсутствует, могут встречаться липидные капли. Остальная часть объема цитоплазмы (около 75%) приходится на воду. У некоторых видов плазмодий содержит много извести (до 28%). Для большинства плазмодиев характерно присутствие пигментов, которые определяют окраску плодового тела, причем она может быть самой разнообразной, но постоянной для конкретного вида. По мнению некоторых ученых, определенные пигменты выполняют роль фоторецепторов, благодаря которым слизевики могут различать освещенные и затемненные места, выбирая их по мере необходимости.
Форма вегетативных тел слизевиков непостоянна и может быть очень разнообразной. Столь же широко варьируют и их размеры, например, у фулиго, или слизевика многоголового, поперечные размеры плазмодия превышают 25 см (у некоторых они могут достигать 1 м в диаметре), тогда как у других они очень маленькие (менее 1 мм).
Большинство слизевиков являются сапрофитами, необходимые вещества их плазмодии поглощают из окружающей среды через всю поверхность. В основном это осуществляется путем обычной диффузии, но они также способны к фагоцитозу твердых пищевых частиц, а также бактерий и простейших (следовательно, они не всегда ведут себя как типичные сапрофиты). Ряд видов слизевиков ведут паразитический образ жизни.
Вегетативная стадия сапрофитных слизевиков тяготеет к сырым и темным местам (т. е. характеризуется положительным гидротаксисом и отрицательным фототаксисом), заселяя упавшие деревья, гнилые пни, или развиваются под опавшей листвой. При наступлении неблагоприятных условий (например, недостаток влаги или низкая температура) плазмодий может уплотняться, затвердевать и переходить в состояние покоящегося склероция (греч. scleros - твердый), в котором может находиться длительное время (многие годы). Когда условия внешней среды станут приемлемыми, из склероция вновь развивается плазмодий.
После определенного периода вегетативного развития и накопления питательных веществ плазмодий переходит к спороношению. Для этого необходим свет, поэтому двигательные реакции меняются на противоположные и плазмодий выбирается на освещенные и более сухие места, причем некоторые виды проявляют склонность к определенному субстрату. Спорообразование сопровождается образованием плодовых тел, внешний вид которых может быть самым разнообразным (рис. 402, 403). В самом простом случае в плодовое тело превращается весь плазмодий, при этом он покрывается оболочкой. У других видов плазмодий распадается на большое количество отдельных спороношений, которые располагаются группами. У некоторых слизевиков отдельные спороношения объединяются в более крупные - эталии, которые покрываются общей оболочкой.
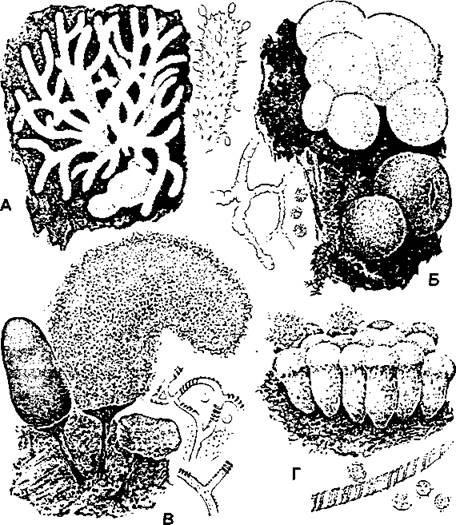
Рис. 402. Слизевики:
А - цератиомикса (Ceratiomyxa): общий вид спороношения, справа - часть спороношения со спорами:
Б - ликовала (Lycogala): молодые (розовые) и старые (бурые) эталии; слева - псевдокапиллиций и споры;
В - арцирия (Агсугіа): внешний вид спороношения с капиллицием (в центре), справа - часть капиллиция и споры;
Г-трихия (Trichia): внешний вид спороношения (внизу - часть капиллиция и споры) (по «Жизнь растений», т. 2)

Рис. 403. Слизевики:
А - дидерма (Diderma): внешний вид спороношения, внизу - часть капиллиция и споры;
Б - стемонитис (Stemonitis): внешний вид спороношения, слева - часть сетчатого капиллиция и споры;
В - физарум (Physarum): общий вид спороношения, внизу - часть капиллиция и споры;
Г-леокарпус (Leocarpus): общий вид спороношения, внизу - часть капиллиция и споры (по «Жизнь растений», т. 2)
В образовавшихся спороношениях развиваются одноклеточные споры, причем спорогенез сопровождается мейотическим делением. Каждая спора покрыта собственной оболочкой, в которой различают два (более тонкий внутренний целлюлозный и наружный хитиновый), а у некоторых видов даже три слоя. У многих видов слизевиков среди спор находятся полые внутри или цельные нити, некоторые из них могут ветвиться. Полагают, что эти нити обладают гигроскопичностью, т. е. способностью обратимо скручиваться и распрямляться под действием поглощенных паров воды. При этом споры разрыхляются. Созревшие гаплоидные споры выходят из спороношения через разрыв в его стенке, подхватываются потоками воздуха и таким образом распространяются. В сухой среде споры слизевиков очень долго (десятки лет) сохраняют жизнеспособность.
Для успешного прорастания споры прежде всего необходима вода. В жидкой среде из споры через имеющиеся поры оболочки или при ее разрыве выходит одна, чаще две, а иногда четыре или даже восемь зооспор, снабженных двумя жгутиками неравной длины (рис. 404). Если спора прорастает не в жидкости, а на увлажненном субстрате, то из нее выходят не зооспоры, а амебовидные организмы, которые называются миксамебами (греч. туха - слизь, amoibe - изменение). И зооспоры и миксамебы размножаются бесполым путем (делением клеток), увеличивая таким образом свою численность.

Рис. 404. Миксомицеты:
А - прорастание споры;
Б - зооспора (по Н. А. Комарницкому и совет.)
После периода бесполого размножения наступает половой процесс. При этом зооспоры или миксамебы сливаются попарно и становятся диплоидными. Интересно, что те гаплоидные клетки, которые не успели прокопулировать, поедаются диплоидными клетками. С диплоидной миксамебы начинается развитие нового плазмодия. Этот процесс сопровождается многократными митотическими делениями ядра, без последующего разделения цитоплазмы на отдельные клетки. Если поблизости оказываются другие диплоидные миксамебы, они также могут сливаться с растущим плазмодием (рис. 405), однако поступившие ядра ни с кем не сливаются, а также делятся митотически. Поступление новых ядер от других миксамеб обеспечивает гетероядерность и генетическое разнообразие.
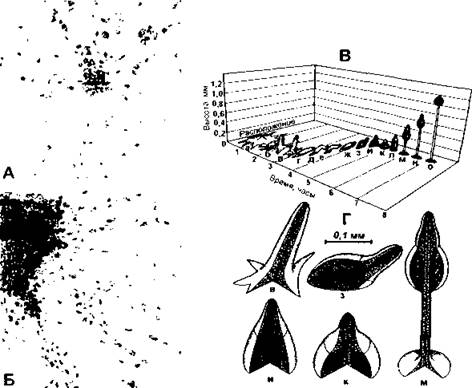
Рис. 405. Размножение миксомицета.
А - отдельные организмы начинают сливаться, образуя плазмодий;
Б - более поздний этап процесса слияния отдельных организмов;
В - схематическое изображение последовательных этапов процесса размножения; а - в - образование плодового тела в результате слияния сотен отдельных амебовидных миксомицетов; г - з - плодовое тело в течение некоторого времени медленно передвигается по поверхности субстрата: и - о- рост стебля, выносящего над поверхностью спорообразующую часть. Процесс приводит к освобождению спор;
Г - отдельные стадии развития плодового тела (по К. Вилли и соавт.)
Формирование плазмодия основано на способности миксамеб к хемотаксису. Как правило, агрегацию отдельных клеток инициирует недостаток питательных веществ. Голодающие миксамебы чутко реагируют на сАМР (цАМФ), который служит внутриклеточным сигналом к объединению клеток. Если вблизи голодающей миксамебы (в любом месте) выделить из микропипетки небольшое количество сАМР, клетка немедленно отреагирует образованием псевдоподии, которая образуется в направлении раздражителя. Установлено, что голодающие миксамебы сами выделяют с АМР, который хемотаксически привлекает других миксамеб. Начало агрегации клеток носит случайный характер - микс- амеба, которая начала первой выделять сАМР, становится центром агрегации миксамеб. Восприятие сАМР осуществляется поверхностными рецепторами клеток. В образовании плазмодия обычно участвуют до 100 000 миксамеб.
Формирующийся вегетативный плазмодий стремится уйти в сырое темное место, где он перемещается в поисках пищи. После завершения вегетативного периода плазмодий приступает к образованию спор и переживает все события, описанные выше.
Среди слизевиков имеются и паразиты высших растений, некоторые из них вызывают заболевания культурных растений, например килу капусты (природу заболевания открыл выдающийся отечественный миколог М. С. Воронин в 70-х гг. XIX в.) или порошистую паршу картофеля (рис. 406). Паразитические слизевики не образуют морфологически отдельных спороношений.
Происхождение слизевиков во многом неясно, полагают, что сапрофитные формы самостоятельно возникли от животных жгутиконосцев в процессе приспособления к наземному существованию. Всего насчитывается свыше 500 видов слизевиков.
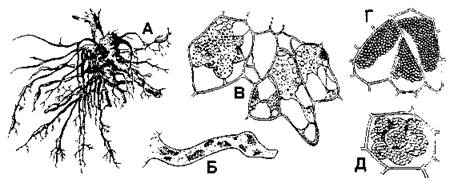
Рис. 406. Кила капусты (А, Б, В, Г) и порошистая парша картофеля (Д):
А - общий вид пораженных килой корней;
Б - вегетативная стадия в корневом волоске;
В - клетки корня с плазмодием возбудителя килы;
Г - клетки корня со спорами паразита;
Д - губчатые комочки спор возбудителя порошистой парши картофеля в тканях клубня (по Т. П. Сизовой (А, В, Г, Д) и Н. А. Комарницкому и соавт. (Б))
Вопросы для самоконтроля и повторения
1. Какова морфология слизевиков?
2. Как осуществляется размножение слизевиков?
3. Как происходит образование плазмодия?
4. Как образуются плодовые тела слизевиков?