Еще в середине XX в. Роберт Эмерсон обнаружил, что если одноклеточную водоросль хлореллу одновременно освещать коротковолновым и длинноволновым красным светом, эффективность фотосинтеза гораздо выше, чем при освещении только одной частью спектра. Это свойство, названное в честь автора эффектом усиления Эмерсона, навело исследователей на мысль, что в хлоропластах одновременно функционируют две взаимосвязанные пигментные системы. Это предположение позднее подтвердилось, когда в мембранах тилакоидов хлоропластов были обнаружены комплексы молекул, названные фотосистемой I и фотосистемой II. Они совместно обеспечивают трансформацию световой энергии в удобную для использования живыми организмами энергию химических связей.
Каждая из фотосистем имеет реакционный центр (рис. 174), который образован пронизывающими насквозь мембрану тилакоида белками, ассоциированными с хлорофиллом (напомним, что комплекс молекулы белка с пигментом называется хромопротеидом). Пигменты реакционного центра способны поглощать энергию света, которая переводит электроны в неустойчивое возбужденное состояние, в результате чего они покидают молекулу хлорофилла и переходят на расположенные поблизости молекулы-переносчики. Это говорит о том, что находящийся в реакционном центре хлорофилл способен осуществлять фотохимические реакции.
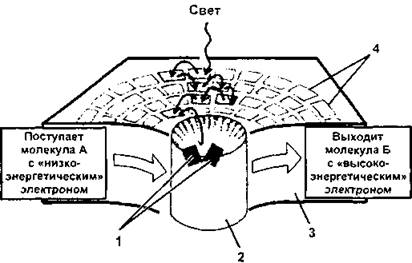
Рис. 174. Схема устройства фотосистемы:
1 - «специальная пара» молекул хлорофилла в реакционном центре; 2 - реакционный центр белково-пигментного комплекса; 3 - мембрана тилакоида; 4 - молекулы хлорофилла в антенном белковом комплексе (по Б. Албертсу и соавт., с изменениями)
Вторым обязательным компонентом фотосистемы является антенный комплекс. В нем также имеется хлорофилл, причем на его долю приходится до 60% общего количества хлорофилла тилакоидных мембран. Специальные исследования показали, что на один реакционный центр приходится 200 - 400 молекул хлорофилла, расположенных в антенных комплексах. Кроме хлорофилла а, здесь присутствуют еще и дополнительные пигменты - хлорофилл в, каротиноиды и фикобилины. Их роль заключается в улавливании света с длиной волн, не доступной для хлорофилла а. Следует отметить, что молекулы пигментов антенных комплексов, пребывая в возбужденном состоянии (в результате поглощения энергии фотона), не осуществляют фотохимических реакций, зато они эффективно передают полученную энергию по цепи хлорофиллу реакционного центра. Направление переноса энергии (электроны здесь не передаются) в антенных комплексах всегда ориентировано от пигментов, поглощающих самую короткую часть спектра (каротиноидов), к более «длинноволновым» пигментам. Как мы уже говорили, такой процесс получил название резонансной передачи энергии. При этом резонансная передача энергии, осуществляемая между одинаковыми молекулами хлорофилла, получила название гомогенной, если же энергия переносится на другой тип пигмента перенос называется гетерогенным.
Рассмотрим устройство фотосистемы I. Считается, что сторически она возникла раньше фотосистемы II и в настоящее время имеется практически у всех фотосинтезирующих организмов, включая способных к фотосинтезу бактерий (у последних отсутствует фотосистема II и, следовательно, не происходит разложения воды и выделения кислорода). В состав реакционного центра этой фотосистемы входят хромопротеиды, содержащие самую длинноволновую форму хлорофилла (его сокращенно обозначают как П700, показывая тем самым длину волны, которую способен поглощать конкретный пигмент). Антенный комплекс этой фотосистемы включает в себя 110 молекул хлорофиллов группы а, имеющих максимумы поглощения от 675 до 695 нм.
Согласно теории эволюции фотосистема II в процессе исторического развития появилась позже. На современном этапе она присутствует у всех зеленых растений, а также у сине-зелёных водорослей. Белковые комплексы фотосистемы II включают в себя несколько более коротковолновые формы хлорофилла. Реакционный центр содержит более коротковолновую форму хлорофилла а - П680. В антенном комплексе имеются хлорофиллы а670-683.
Кроме того, в мембранах тилакоидов находятся непосредственно связанные с фотосистемой П светособирающие белковые комплексы, в которых присутствуют хлорофиллы a660-675, а также хлорофилл в650 (несколько в большем количестве, чем хлорофилл а) и каротиноиды.
Весьма сложные и разнообразные реакции фотосинтеза, в основе которых лежат фотохимические процессы, в конечном итоге преобразуют энергию света в химическую. Однако наличие света необходимо отнюдь не для всех этапов, а лишь вначале, поэтому в фотосинтезе выделяют световую и темповую стадии.