Благодаря тому, что процесс микроэволюции, завершающийся видообразованием, протекает на протяжении всего существования жизни на Земле, в живой природе образовались естественные группировки видов разного ранга родства, называемые таксонами. Близкородственные виды входят в состав одного рода, родственные роды объединяются в семейства, далее следуют отряды, классы и типы. Тип - наиболее крупный таксон, объединяющий виды, которые имеют сходный общий план строения благодаря отдаленному родству (т.е. происхождению от одного или немногих общих отдаленных предков).
Процесс формирования таксонов надвидового ранга называют макроэволюцией. Процесс макроэволюции изучают методами палеонтологии, сравнительной анатомии, сравнительной эмбриологии, а также многих других современных наук: генетики, биохимии, иммунологии, физиологии, биогеографии и др.
13.1.1. Темпы эволюционных преобразований
Синтетическая теория эволюции, основные положения которой подробно изложены в предыдущих главах, сложилась в 30-40-х гг. прошлого века в результате объединения теории Дарвина с классической генетикой (в первую очередь речь идет о популяционной генетике). За прошедшие с тех пор годы получены новые фактические материалы в области палеонтологии, сравнительной и экспериментальной биологии, значительные успехи достигнуты в молекулярной генетике, в частности, прочитаны геномные тексты различных про- и эукариотических организмов. Все эти данные значительно дополнили и обогатили представления об эволюционном процессе, его закономерностях и механизмах, позволили внести ясность в решение ряда вопросов, которые дискутировались учеными-эволюционистами на протяжении всего времени существования теории.
Один из таких вопросов - происходит ли эволюция плавно и постепенно - градуалистически - или неравномерно (скачками), как утверждали приверженцы сальтационизма (рис. 13.1)? Замечено, что темпы эволюции, которые можно охарактеризовать появлением новых таксонов, колеблются в широких пределах. Известно, что все типы беспозвоночных появились в относительно короткий период на рубеже протерозойской и палеозойской эр, причем в кембрийском периоде типов было больше, чем сейчас, - некоторые тогда же и вымерли. Хордовые возникли несколько позднее, но тоже в течение кембрия. Столь же высокие темпы формирования крупных таксонов демонстрируют высшие растения в начале освоения суши - все отделы (соответствуют типам животных) споровых и голосеменные появились в девонском периоде. Так же быстро происходило формирование систематических групп млекопитающих и цветковых после мелового кризиса. На основании этих и аналогичных фактов многие палеонтологи независимо друг от друга пришли к мысли о неравномерности эволюционных преобразований. Применение методики количественной оценки темпов эволюции на основе сравнения нуклеотидных последовательностей ДНК различных групп организмов, позволило реконструировать их эволюционное развитие и разрешить давний спор между приверженцами градуализма и сторонниками скачкообразной эволюции. Проанализировав «эволюционные древа», построенные для 122 групп организмов, ученые пришли к выводу, что в среднем 22% различий в ДНК возникает во время кратких периодов интенсивного видообразования, а остальные 78% постепенно накапливаются во время долгих «градуалистических» периодов. Неравномерность эволюции в большей мере свойственна растениям и грибам и в меньшей - животным.

Рис. 13.1. Модели эволюции: а - градуализм; б - сальтационизм
Что же становится причиной всплеска эволюционных преобразований? Многие эволюционисты считают основной причиной изменения, происходящие в биоценозе (экосистеме), которые могут быть обусловлены, в том числе, биосферными кризисами.
Организмы существуют не сами по себе, а в составе сложных систем - сообществ, или биоценозов. В сложившемся сообществе виды тесно взаимно адаптированы. Все ниши заняты, свободных мест нет или мало. Виды, потенциально претендующие на одни и те же ниши (чаще всего это близкородственные виды, входящие в один старший таксон) стремятся разделить экологическое пространство, чтобы ослабить конкуренцию. Сужение ниши дает возможность лучше к ней приспособиться, повысить эффективность использования ресурсов. Таким образом, виды продолжают изменяться, и эти изменения направляются и регулируются сообществом. Важнейшие направления эволюции видов в устойчивом биоценозе - экологическая специализация и коадап-тация (взаимная адаптация), т.е. происходит согласованная - когерентная - эволюция (коэволюция).
Устойчивое сообщество способствует экологической специализации еще и тем, что оно в значительной степени повышает стабильность условий существования для входящих в него видов. Так как соседи по биоценозу, составляющие важнейший элемент внешней среды для каждого вида, в основном одни и те же, к ним легко приспособиться, как к постоянным факторам среды. Кроме того, живые организмы стабилизируют и иные параметры среды, например, каждый устойчивый тип растительного сообщества создает и поддерживает определенный состав почвы, затененность, влажность, свой микроклимат.
Ситуация в корне меняется во время экологических кризисов, когда структура сообществ разрушается и экосистемная регуляция эволюции временно ослабевает. В эти кризисные периоды эволюция организмов становится более быстрой и несогласованной - некогерентной.
При разрушении биогеоценоза большая часть видов погибает вместе с ним, причем в первую очередь вымирают самые специализированные, т.е. самые приспособленные и конкурентоспособные в недавнем прошлом. При этом освобождается множество ниш. Стабилизирующий отбор, удерживавший каждый вид в строгих рамках, определявшихся структурой сообщества, резко ослабевает. Растет изменчивость и наблюдается взрыв формообразования - все освободившиеся ниши занимают иные виды. Это продолжается до тех пор, пока экологическое пространство снова не заполнится и не произойдет организации возросшего разнообразия видов в новые структурированные сообщества, которые будут уже другими, и направленность эволюционных процессов, задаваемая новой структурой биоценозов, сможет отличаться от прежней при сохранении основных тенденций - развития в направлении специализации и коадаптации.
Примеры подобных скачкообразных эволюционных преобразований при разрушении экосистемы наблюдались в течение последних десятков лет в Аральском море. В 60-е годы прошлого века воду из рек, наполняющих это море, стали забирать на орошение полей. В результате площадь моря начала резко уменьшаться и теперь Арала как единого водоема уже не существует: он разделился на два изолированных, быстро высыхающих озера - Большой и Малый Арал (рис. 13.2).
В обоих водоемах резко выросла соленость, что привело к вымиранию большей части видов животных и растений. Массовое вымирание освободило множество ниш. Например, вымерли все моллюски-фильтраторы, однако многие двустворчатые моллюски - зарывающиеся грунтоеды - уцелели. Резкая перемена условий привела к тому, что выжившие виды начали быстро эволюционировать. У них резко возросла изменчивость и появились целые «пучки» новых форм. Различия между этими новыми формами и исходными видами порой очень велики. Такая степень различий характерна для разных родов, а иногда и семейств двустворчатых моллюсков. Пустующие ниши стали заселяться выжившими моллюсками, которые ранее вели совсем другой образ жизни. В результате зарывающиеся грунтоеды из рода Cerastoderma стали перемещаться на поверхность грунта и превращаться в фильтраторов, при этом происходили соответствующие изменения в строении их раковины. Все эти эволюционные процессы протекали в кратчайшие сроки и очень сходным образом в двух разобщенных водоемах - Большом и Малом Арале.

Рис. 13-2. Снимок территории Аральского моря со спутника: а - сентябрь 1989 г.; б - октябрь 2008 г.
Так как биогеоценоз, как любая система, представляет собой совокупность взаимосвязанных и взаимодействующих элементов, то совершенно очевидно, что не только сообщество влияет на эволюцию видов, но и изменяющиеся виды тоже оказывают воздействие на экосистемы. Так, например, трехиглая колюшка - небольшая рыбка, широко распространенная в морях, реках и озерах Северного полушария, около 10 000 лет назад начала осваивать пресные водоемы, что было связано с отступлением ледников и образованием на их месте множества больших и малых озер. Это стало стимулом для быстрых эволюционных изменений у колюшки, которые оказались наиболее выраженными в семи озерах у тихоокеанского побережья Юго-Западной Канады, где эта рыба фактически разделилась на два самостоятельных вида: бентосный, т.е. донный, и пелагический, живущий в толще воды. Первый вид питается донными беспозвоночными, второй охотится на планктонных беспозвоночных в верхних слоях воды. Виды различаются не только диетой и образом жизни, но и размерами (донный вид существенно крупнее), формой тела, некоторыми деталями строения, а также брачным поведением. Виды практически не скрещиваются между собой. Выполненные в этих озерах исследования показали, что видообразование у колюшки заметно влияет на озерные экосистемы, в том числе на прозрачность воды, состав растворенной органики, количество планктона и относительную численность многих видов мелких животных. Очевидно, что изменение свойств экосистемы, в свою очередь, должно влиять на эволюцию населяющих ее видов. Таким образом, эволюционные изменения, затронувшие один вид организмов, могут иметь далеко идущие последствия для многих других видов.
Количественный анализ динамики видового разнообразия в биоценозах подтверждает существование длительных периодов сравнительно медленных, коадаптивных изменений, сменяемых кратковременными кризисами, во время которых структура сообществ разрушается, и происходят быстрые, малопредсказуемые эволюционные процессы. Отмечается, что рост биоразнообразия в периоды интенсивного видообразования в развивающейся экосистеме может быть самоускоряющимся, т.е. видообразование у одних групп организмов стимулируют этот процесс и у других групп организмов. Складывающаяся после кризиса новая структура сообществ в большинстве случаев оказывается более сложной и более устойчивой, чем прежняя.
13.1.2. Механизмы эволюционных преобразований
Достижения молекулярной генетики, активно развивающейся последние десятилетия, позволили значительно расширить представления о возможных путях эволюционных преобразований организмов в составе таксонов различного ранга - от вида до типа. Свой вклад внесли прочтение текстов ДНК различных групп организмов, развитие представлений о системной организации генома, обнаружение ряда генов и установление их гомологии и функций в разных таксономических группах, изучение молекулярных механизмов, контролирующих функцию генов и обеспечивающих формирование фенотипических признаков организмов, открытие мобильных генетических элементов и многое другое.
В соответствии с основными положениями синтетической теории эволюции, основную роль играют генные мутации, на основе которых формируются новые признаки, подвергаемые естественному отбору. И действительно, такой механизм имеет место, что также было подтверждено современными молекулярно-генетическими методами. При сравнении структуры антител, синтезируемых организмом при встрече с новым штаммом возбудителя, ученые обнаружили, что больше всего мутаций происходит в трех небольших участках длинной молекулы. Сложнейшими экспериментами было доказано, что мутированию подвергаются не только эти участки гена, но и другие, причем мутации везде происходят с одинаковой скоростью. Однако только мутации, произошедшие в трех определенных участках молекулы, могут улучшить связывание возбудителя. Каждое новое антитело испытывается организмом, и все не подходящие к данному возбудителю отбраковываются. Таким образом, сохранение только отдельных мутаций из всего спектра представляет собой результат отбора.
Однако, как установлено к настоящему времени, эволюция на основе мутаций кодирующих последовательностей структурных генов - только один из механизмов. Другие возможные пути эволюционных преобразований - изменение некодирующих регуляторных последовательностей ДНК, дупликации генов, изменения, связанные с перемещением мобильных генетических элементов, перенос генетической информации от одного организма к другому (возможно, другого вида), хромосомные мутации, гибридизация (возможно, межвидовая), наследуемые изменения экспрессии генетической информации.
Установлено, что значительная часть нуклеотидных последовательностей в геноме эукариотических организмов некодирующая, причем в ходе эволюции эукариот доля этой части в целом возрастает. Так, в геноме человека кодирующими являются чуть более 1% нуклеотидных последовательностей ДНК. Обнаружено также, что большинство мутаций происходит не в них, а в регуляторных областях молекул ДНК. Сравнение геномов сумчатых млекопитающих и плацентарных показало, что за 180 млн лет, прошедших после разделения этих линий, в кодирующих белок областях геномов возникло сравнительно мало мутаций. Ведущая роль в эволюции высших организмов принадлежит изменению регуляторных участков генома, которые сами белки не кодируют, но влияют на работу кодирующих белок генов. Пример подобных изменений обнаружен при исследовании гена Duffy человека, кодирующего поверхностный рецептор, взаимодействие с которым необходимо для проникновения малярийного плазмодия в клетку. В аборигенных популяциях Африки в регуляторном районе этого гена зафиксирована мутация - замена тимина на цитозин, повреждающая сайт связывания транскрипционного фактора GATA. В результате этого транскрипции гена не происходит, и африканцы не имеют антигена Duffy, что обеспечивает устойчивость аборигенов Африки к малярийному плазмодию. У большинства белых европейцев регуляторный сайт гена не поврежден, что делает их восприимчивыми к малярии.
Значение подобного типа мутаций продемонстрировано и в экспериментах. В одном из них регуляторную область гена летучей мыши пересадили обычной мыши. Ген контролирует время пролиферации хрящевых клеток передней конечности до начала их превращения в клетки кости. Пересадка регуляторной области привела к удлинению передних лап животного-реципиента на 15%.
Еще одним открытием, важным для понимания реализации эволюционных преобразований, стало установление огромной роли мобильных генетических элементов в возникновении мутаций. МГЭ открыты американским генетиком Б. МакКлинток в 1951 г. Это последовательности, наличие которых приводит к нестабильности генома. Они способны активно размножаться и перемещаться с места на место в ДНК. При размножении они передаются в ряду поколений - от родителей к потомкам, а некоторые из них могут передаваться и горизонтально, между таксонами. Многие МГЭ многократно повторяются в геноме. Перемещение МГЭ может вызвать мутации кодирующей части генов, регуляторных последовательностей и т.д. Наличие большого числа копий МГЭ в геноме резко повышает вероятность ошибок в ходе репликации и рекомбинации ДНК, что также приводит к мутациям.
В геноме дрозофилы обнаружено более 30 семейств МГЭ, которые занимают примерно 15% генома. Установлено, что у D. melanogaster 70% мутаций вызваны встраиванием МГЭ.
Геном человека почти наполовину (45,5%) состоит из всевозможных МГЭ - транспозонов, ретротранспозонов и т.п. У других организмов их объем также довольно значителен: у мыши - 40,9%, у опоссума - 52,2%, у курицы - 9,4% генома. У прокариот мобильных элементов меньше, но в их геномах МГЭ также присутствуют.
Перемещаясь, МГЭ могут перетаскивать с собой небольшие фрагменты ДНК, встраивая их в новое место той же или другой молекулы. В ряде случаев это приводит к созданию новых кодирующих последовательностей ДНК, возникновению новых регуляторных элементов, дуплицированию (удвоению) целых генов.
Так, обнаружено, что в ходе эволюции у приматов появилось много новых генов, причем в основном в результате удвоения старых. Эволюция одного из таких генов, CDC14Bretro, исследована очень подробно. Установлено, что он появился у общего предка человекообразных обезьян в результате деятельности МГЭ - ретротранспозонов. Позднее, у общего предка гориллы, шимпанзе и человека, ген претерпел быстрые изменения под действием отбора, сменив свою локализацию в клетке и функцию. Изначальный вариант белка занимался отрезанием фосфатных групп у некоего вещества в центре организации микротрубочек и тем самым управлял клеточным делением. Примерно то же самое первоначально делал и белок, кодируемый вновь образовавшимся ретрогеном CDC14Bretro, но уже не во всех тканях, а только в мозге и семенниках. У гиббонов и орангутанов он сохранил эту функцию до сих пор. Затем 14-7 млн лет назад у предков африканских человекообразных обезьян ретроген попал под действие отбора и быстро накопил 12 значимых замен, которые привели к смене внутриклеточной локализации и функции белка. Теперь белок стал присоединяться к мембранам эндоплазматической сети и отрезать фосфатные группы у какого- то другого вещества (об этом свидетельствует изменение активного центра).
Считают, что дупликации генов с последующим расхождением их функций - главный путь эволюционных преобразований. Подобным образом сформировалось огромное число генов в различных группах живых организмов, например, гены, кодирующие белки-кристаллины хрусталика, появившиеся у древнейших позвоночных животных, или гены, кодирующие белки- антифризы, препятствующие замерзанию воды в клетках рыб, обитающих в районе полюсов Земли.
Сравнительный анализ геномов сумчатых и плацентарных млекопитающих обнаружил, что с момента их расхождения (дивергенции) 15% генов в линии сумчатых (опоссума) и 20% в линии человека подверглись одной или нескольким дупликациям. В основном это гены «экологического» характера, отвечающие за взаимодействие с внешней средой: вкусовые и обонятельные рецепторы, гены иммунной системы, пищеварительных ферментов, а также гены белков, отвечающих за обезвреживание токсичных веществ (например, цитохром P450). Формирование некодирующих последовательностей в геномах организмов в ходе эволюции также осуществлялось при участии МГЭ. Расшифровка и сравнение геномов ряда млекопитающих (человека, мыши, крысы, собаки, опоссума) показало, что значительная часть появившихся у этих животных новых некодирующих последовательностей, выполняющих регуляторные функции, сформировалась из фрагментов МГЭ - транспозонов.
Помимо существенного вклада МГЭ в процессы, описанные выше, эти элементы активно участвуют в горизонтальном переносе генетического материала, т.е. передаче генетического материала между одновременно живущими особями, причем не обязательно одного вида. Значение этого процесса для эволюции чрезвычайно велико.
Так, оказалось, что гены и даже целые геномы паразитических бактерий иногда могут вставляться в хромосомы животных-хозяев. В геномах 4 видов насекомых и 4 видов круглых червей обнаружены фрагменты генома внутриклеточной паразитической бактерии Wohlbachia (см. п. 2.4.3.4). В ряде случаев, например, у тропической плодовой мушки Drosophila ananassae, геном бактерии оказался вставлен в геном хозяина практически целиком, при этом многие гены, заимствованные мухой у бактерии, активно работают. Таким образом, внедрение ДНК организма другого таксона может быть одним из способов приобретения новых генов и признаков в эволюции высших организмов. Чаще всего перемещаются вирусные и бактериальные ДНК.
Наибольшее значение горизонтальный перенос генетического материала имеет в эволюции простейших форм жизни. Так, в геноме у одной из групп диатомовых водорослей Phaeodactylum обнаружилось 587 генов, заимствованных у различных прокариот: цианобактерий, протео-бактерий, архебактерий и других. 170 генов они получили от красных водорослей - эукариотических организмов.
Направленное изучение явления горизонтального обмена генами в разных таксономических группах организмов установило, что развитие органического мира не похоже на эволюционное древо, оно представляет собой сплетение или сеть (рис. 13.3).
Эволюционные преобразования возможны не только вследствие приобретений, но и потерь, т.е. утрата генов может открывать путь для прогрессивных изменений. Недавно обнаружилось, что некоторые гены, которые нормально работают в организме шимпанзе, у человека выключились и превратились в молчащие «псевдогены». Например, выключение гена MYH16 привело к уменьшению (редукции) жевательной мускулатуры у предков рода Homo, а это, в свою очередь, позволило мозгу начать увеличиваться. В геноме человека выявлено еще около 50 молчащих генов, аналоги которых у шимпанзе нормально функционируют. Работают они и у других обезьян, фрагменты генома которых уже прочтены.
В последние годы все более пристальное внимание эволюционисты обращают на роль хромосомных и геномных мутаций в эволюции. Считают, что наибольшее значение среди хромосомных перестроек имеют, прежде всего, инверсии и транслокации. Примеров таких мутаций достаточно много. Так, известно, что геномы человека и шимпанзе различаются десятью крупными геномными перестройками: одним слиянием и девятью инверсиями. Еще один яркий пример роли хромосомных перестроек - формирование целой группы таксонов в надвиде слепушонка Ellobius talpinus. На рис. 13.4 представлен так называемый «робертсоновский веер» - хромосомные перестройки путем слияния акроцентрических хромосом.

Рис. 13.3. Вклад горизонтального переноса генетического материала между таксонами в эволюцию

Рис. 13.4. Робертсоновский веер у слепушонок надвида Ellobius talpinus (по: Воронцов Н.Н., 1980)
Полиплоидия - также довольно распространенный механизм эволюции, причем встречается она не только у растений, но также и в других царствах. В последние годы полиплоидия открыта у целого ряда животных - рыб, амфибий, рептилий. Установлен факт, что на заре эволюции позвоночных одна за другой произошли две полногеномные дупликации, что привело к учетверению всего генома.
Определенный вклад в эволюционный процесс вносит и межвидовая гибридизация. Иллюстрацией может служить ситуация в зоне симпатрического обитания двух близких видов - обыкновенной и бело-шапочной овсянок - около 2,5 тыс. км с запада на восток, от Уральских гор до Байкала. Здесь наблюдается весьма высокий уровень гибридизации, доля гибридов в отдельных популяциях достигает 40-45%. Процесс гибридизации динамично развивается во времени: для двух популяций за 30 последних лет прослежено резкое снижение численности белошапочной овсянки, вплоть до ее полного исчезновения, и увеличение доли фенотипических гибридов.
Активно обсуждается вклад в эволюционный процесс открытого в конце 1970-х гг. явления кардинальной пространственной реорганизации наследственного материала. Этот феномен был обнаружен при изучении распределения хроматина в ядрах клеток малярийного комара. Установлены существенные различия в трехмерной организации хромосом между разными, в том числе и очень близкими, видами. Видо-специфичен ряд характеристик, среди которых наличие-отсутствие связей хромосом с ядерной оболочкой, морфология хромосомных участков прикрепления, локализация мест контактов на хромосомах.
Аналогичные данные получены и при изучении пространственной локализации хроматина в ядрах клеток дрозофилы. Показано, что изменение архитектуры интерфазных ядер может рассматриваться как механизм эволюционных преобразований таксонов.
На сегодняшний день хорошо известно, что не вся наследственная информация, передающаяся из поколения в поколение, кодируется последовательностью нуклеотидов молекул ДНК. Кроме этой генетической информации есть еще и так называемая эпигенетическая. Эпи-генетика («эпи» - в переводе с греческого «над») - раздел современной биологии, предметом которого является изучение наследования в ряду клеточных и организменных поколений функциональной активности генов, не связанное с изменением первичной структуры входящей в их состав ДНК.
К такого рода эпигенетическим явлениям относят, в том числе, пара-мутации, РНК- интерференцию, метилирование ДНК.
Парамутация - это устойчивое, наследуемое изменение свойств аллеля гена, возникающее в результате взаимодействия его с другим аллелем того же гена, при этом нуклеотидная последовательность ДНК не меняется. Парамутации впервые обнаружены у растений полвека назад, а недавно они зарегистрированы и у животных.
Классический пример парамутации дает один из аллелей гена pl1 - Pl1-Rhoades, от которого зависит окраска пыльников у кукурузы (рис. 135). Ген pl1 кодирует белок - транскрипционный фактор, регулирующий активность ряда других генов, которые, в свою очередь, кодируют белки, необходимые для синтеза пигмента антоциана. Аллель Pll-Rhoades при одной и той же последовательности нуклеотидов может находиться в одном из двух состояний: «активном» (Pl- Rh, пурпурные пыльники) и «неактивном» (Pl, светлые пыльники). Суть явления в том, что неактивное состояние аллеля Pl как бы «заразно», то есть аллель в активном состоянии Pl- Rh, побывав в одной клетке с неактивным аллелем Pl', сам превращается в Pl.

Рис. 13.5. Фенотипическое проявление парамутации аллеля Pl1-Rhoades. Слева - пурпурные пыльники (ген в «активном» состоянии, Pl-Rh), справа - светлые пыльники (ген в «пассивном» состоянии, Pl')
На сегодняшний день удалось выяснить, что большую роль в поддержании «переключенного» состояния гена при парамутации играют молекулы РНК, передающиеся от родителей к потомкам. Кроме того, недавно выяснилось, что для устойчивой передачи парамутации из поколения в поколение необходим фермент, размножающий молекулы РНК без участия ДНК - РНК-зависимая РНК-полимераза.
С участием РНК реализуется еще один эпигенетический феномен - РНК-интерференция (см. п. 8.2.5.2). Это механизм подавления экспрессии гена на стадии трансляции. Комплементарное соединение малой интерферирующей РНК с мРНК, подлежащей трансляции, приводит к разрезанию последней ферментами (нуклеазами), и белок не синтезируется.
В 2002 г. исследователи неожиданно выяснили, что действие малых интерферирующих РНК может не ограничиваться только лишь временным выключением генов на уровне мРНК. Имеются данные, что в некоторых случаях эта РНК, видимо, воздействует прямо на ДНК, изменяя структуру хроматина и способствуя длительному молчанию одних, и, возможно, активизации других генов, что, безусловно, можно рассматривать как один из возможных эволюционных механизмов. Явление РНК-интерференции лежит, например, в основе разнообразия окраски растений петунии, имеющих один и тот же исходный генотип, при введении трансгена с интерферирующей РНК (рис. 13.6).
Другое эпигенетическое изменение экспрессии гена - метилирование ДНК (см. п. 8.2.5.2). Метилирование ДНК считается присущим, в основном, эукариотам, и заключается в присоединении метильной группы к цитозину в составе динуклеотида ЦГ. Около 60-70% всех ЦГ- динуклеотидов у млекопитающих метилированы. У человека метилировано около 1% геномной ДНК. Репрессия транскрипции в этом случае опосредована метилцитозин-связывающими белками, которые способны связываться с метилированными ГЦ- динуклеотидами. Они, в свою очередь, привлекают деацетилазу гистонов и другие факторы, участвующие в модификации гистонов. В результате формируется конденсированная транскрипционно неактивная структура гетерохроматина. Метилирование ДНК связано также с такими феноменами, как геномный импринтинг и инактивация Х-хромосомы.

Рис. 13-6. Пример РНК-интерференции при получении трансгенных растений петунии. Крайний слева цветок - дикий тип, два других цветка содержат трансген, РНК которого включает РНК- интерференцию и в результате подавляет выражение гена, ответственного за синтез пигмента
Влияние метилирования ДНК на структуру хроматина имеет большое значение для развития и функционирования живого организма. В частности, отсутствие метилцитозин-связывающего белка 2 (MeCP2) приводит у человека к формированию синдрома Ретта - заболевания, характеризующегося психоневрологическими нарушениями и тяжелой умственной отсталостью.
Считают, что эффекты метилирования, приводящие к преобразованию структуры хроматина и, вследствие этого изменяющие активность генов, также играют определенную роль в эволюции организмов. Установлено, что количество метилированных остатков цитозина возрастает с увеличением размера генома и сложности организма.
Помимо выявления разнообразных возможных механизмов эволюционных преобразований исследования последних лет показали, что мутирование разных участков ДНК идет с неодинаковой скоростью, мутации могут носить относительно направленный характер, эволюционные изменения могут единовременно затрагивать совокупность признаков и наблюдаться не у отдельной особи, а одновременно в целой группе организмов.
Ученые, работающие с геномами ранних зародышей дрозофил, обнаружили новый тип регуляторных нуклеотидных последовательностей - теневые энхансеры. Их действие такое же, как и основных (см. пп. 2.4.5.5-А, 8.2.5.2), только обнаружить его можно лишь при отключении основных энхансеров. Установлено, что теневые энхансеры мутируют в среднем в 1,6 раз быстрее, чем основные.
Это не единственный пример последовательностей, имеющих повышенную скорость мутаций. Так, в геноме человека выявлен ряд последовательностей, мутации в которых происходят чаще, чем в среднем. Считают, что именно эти последовательности играли особенно важную роль в становлении человека.
Интересный результат был получен при сравнении более 3000 пар ортологичных генов человека и мыши. Ортологичными называют гены в геномах организмов разных таксонов, исходно произошедшие от одного предкового. Оказалось, что чем больше синонимичных, т.е. не ведущих к изменению структуры кодируемого белка, мутаций произошло в гене за длительное время, тем большее в нем число несинонимичных (значимых) мутаций.
Выше уже обсуждалось, что на частоту возникновения мутаций влияют и перемещения МГЭ. При встраивании их в определенные локусы частота мутирования этих локусов может превышать обычную в тысячи раз.
Многие МГЭ (транспозоны) способны встраиваться не в любое место генома, а только туда, где есть определенная последовательность нуклеотидов. Таким образом, мутации, вызываемые подобными перестройками, не совсем случайны. Все это придает эволюционным изменениям, происходящим при участии МГЭ, отчасти направленный характер. Кроме того, известно, что мутации ряда последовательностей ДНК, в частности, кодирующих р-РНК, т-РНК, гистоны, в большинстве своем являются «запрещенными». Это делает невозможным эволюционные изменения в данных направлениях.
Возможность изменения целой группы признаков даже при единичной точечной мутации определяется наличием в геномах организмов регуляторных последовательностей и генов- регуляторов. Так, мутация, затрагивающая ген-регулятор, кодирующий, например, транскрипционный фактор, приводит к одновременному изменению работы всего комплекса контролируемых им генов и, как следствие, к изменению множества определяемых ими фенотипических признаков. Например, при дифференцировке эритроцита регуляторный фактор GATA1 активирует группу генов, кодирующих все ферменты, обеспечивающие биосинтез гема, а также и а- и β-субъединицы гемоглобина. По выполненным расчетам, вероятность того, что множественные изменения возникнут в результате независимого возникновения мутаций по всей этой группе генов одновременно, практически равна нулю.
Изменение регуляторных последовательностей часто наблюдается при перемещении МГЭ. В эксперименте с арабидопсисом подобным единичным перемещением удавалось получить качественные изменения морфологии растения. Например, возможно возникновение карликовых растений с ранним цветением, извитыми листьями и терминальным цветком. Возможен цветок из двух рядов лепестков и тычинок, в котором чашелистики замещаются лепестками, а пестик тычинками и т.д. Геномы всех полученных растений практически идентичны, так как отличаются лишь небольшими фрагментами ДНК. При этом растения характеризуются принципиальными фенотипическими различиями, которые требуют отнесения их к различным таксонам.
Сравнение геномов опоссума и человека обнаружило, что значительное количество встраиваний МГЭ, произошедшее с момента дивергенции этих групп организмов, случилось в окрестностях 240 генов, кодирующих ключевые регуляторы индивидуального развития - НОХ- генов (см. п. 8.2.10.1). Предполагают, что следствием этого стало изменение регуляции этих генов, послужившее причиной преобразований в строении организма у плацентарных млекопитающих. Модификации регуляторных последовательностей могут приводить к изменению места и времени активности подобных генов, что может проявляться в формировании структур в нетипичных местах организма, изменении протекания стадий онтогенеза, например ускоренном половом созревании или замедлении соматического роста у животных. Аналогичные последствия могут иметь и точечные мутации в самих этих генах. Параметры экспрессии гена зависят, кроме того, от функционального состояния хроматина рядом с ним. Из этого следует предположение, что хромосомные перестройки, вероятно, также могут модифицировать временные и пространственные параметры экспрессии генов.
Ряд авторов полагает возможным осуществление описанных изменений в предшественниках половых клеток до вступления в мейоз. Вследствие этого появляется сразу группа особей с измененными характеристиками, что, возможно, приводит к их репродуктивной изоляции. Показано, что массовые фенотипические изменения могут происходить при активизации перемещений МГЭ в условиях дестабилизации (разрушения) экосистем.
Интенсивность исследований в этой области стремительно нарастает в последние десятилетия и в ближайшие годы можно ожидать новых открытий.
13.1.3. Принципы эволюционных преобразований
В настоящее время считается, что многие, возможно, даже все основные, эволюционные новообразования появились не в результате постепенного накопления мутаций, а за счет перекомбинации готовых функциональных блоков живых систем. Такими блоками в разные моменты эволюционного развития могли служить молекулы, гены, клетки, структуры организма, целые особи и их сообщества. Так формулируется блочный (модульный) принцип в теории эволюции.
Как отмечалось, еще до появления жизни, в ходе абиогенного синтеза, вполне могли сформироваться короткие белковые молекулы, представляющие собой случайные комбинации аминокислот (см. п. 1.4.2). Оказалось, что уже такие короткие белки обладают слабыми каталитическими свойствами, причем различающимися у разных молекул. Большие сложные белки могли образоваться как комбинации из таких сравнительно коротких кусочков-блоков. Анализ структуры известных белков подтверждает именно такой путь развития.
Первые живые системы, по одной из гипотез, возникли путем объединения абиогенно синтезированных РНК и белковых молекул.
Формирование новых генов и других функциональных последовательностей ДНК происходит при перетасовке генетических модулей. Хромосомные перестройки - образование новых комбинаций предсуществующих блоков - также не редкость среди механизмов эволюционных преобразований. Иллюстрация реализации блочного принципа - наблюдаемое повсеместно явление дупликации генов с возможным последующим расхождением их функций. Горизонтальный перенос генетического материала - перемещение в геном организма МГЭ, генов и целых геномов других организмов - также пример блочного характера эволюции. Блочный принцип эволюционных преобразований проявляется и в феномене симбиоза. Существует мнение, что появление новых форм жизни в результате симбиоза, то есть при слиянии сообщества или каких-то его компонентов в единый организм, - один из основных путей эволюции.
Так, эукариотическая клетка возникла в результате симбиоза нескольких разных видов прокариот-бактерий. Эти бактерии сначала долго существовали как компоненты интегрированного бактериального сообщества - бактериального мата (см. п. 1.4.4) (рис. 13.7). После того как между ними установилась устойчивая система взаимодействий и взаимной координации, эти бактерии слились в единый организм, который и стал первой эукариотической клеткой. Хорошо известен пример с лишайниками - организмами, которые представляют собой сожительство гриба с одноклеточными водорослями. Лишайники, по сути, представляют собой маленькие компактные сообщества, которые сами обеспечивают себя всем необходимым. Еще пример - глубоководные рыбы, светящийся орган которых заселен светящимися бактериями. Жвачные млекопитающие никогда бы не научились питаться грубой растительной пищей, а значит, и не стали бы жвачными, если бы не обзавелись ценными сожителями - одноклеточными симбионтами, живущими у них в желудке и переваривающими за них целлюлозу, которую ни одно млекопитающее переварить самостоятельно физиологически не может. Термиты, которые питаются исключительно древесиной, для разложения целлюлозы имеют в своем кишечнике целую группу симбионтов - жгутиконосцев и живущих внутри них бактерий. Сначала термиты находят пищу и измельчают древесную массу, доводя до мелкодисперсного состояния, в котором ее могут поглощать жгутиконосцы. Затем за дело берутся живущие внутри жгутиконосца бактерии. Наряду с генами, отвечающими за синтез целлюлазы - фермента, разрушающего молекулы целлюлозы, в их геноме есть гены, кодирующие ферменты, ответственные за азотфиксацию, - связывание свободного азота атмосферы N2 и превращение его в форму, пригодную для использования не только самими бактериями, но также жгутиконосцами и термитами. Более того, термиты не только сами используют, но и при необходимости «заражают» друг друга этими симбионтами. Значительную роль играет блочный принцип эволюции и при формировании ароморфозов, хотя это проявляется и не столь ярко. Ароморфоз - появление прогрессивных крупных качественно новых эволюционных изменений у организмов, приводящих к повышению их уровня организации и общей энергии жизнедеятельности, позволяющих существовать в более разнообразных условиях среды по сравнению с их предками. Именно появление ароморфозов лежит в основе формирования крупных таксонов надвидового ранга, таких как класс, тип.

Рис. 13.7. Мат из хемосинтезирующих бактерий вблизи гидротермального источника на морском дне
В последние десятилетия существенно изменились представления о том, каким образом происходили в ходе эволюции эти крупнейшие прогрессивные изменения. Оказалось, что во многих случаях переход на более высокий уровень организации происходит не в какой-то одной эволюционной линии, а в нескольких, развивающихся параллельно. При этом отдельные признаки, из которых складывается ароморфоз (блоки), иногда появляются в разных линиях почти одновременно, а иногда - в разное время и в разном порядке. Развивается довольно большое многообразие переходных форм, у которых прогрессивные признаки распределены мозаично. Эти признаки постепенно накапливаются, пока, наконец, в одной или немногих эволюционных линиях они не соберутся все вместе. Для большинства же линий появившиеся отдельные прогрессивные признаки оказываются малозначимыми, и эти линии быстро вымирают. Таким образом, ароморфоз - не единичное эволюционное событие, а довольно сложный комплексный процесс, захватывающий сразу несколько разных родственных групп организмов.
Сейчас многие палеонтологи пришли к выводу, что независимое параллельное приобретение сходных признаков в разных родственных группах организмов при становлении нового крупного таксона - скорее правило, чем исключение. Этот принцип эволюции получил название параллелизма.
Так, всем известен археоптерикс - существо, сочетающее признаки хищных динозавров и птиц. Долгое время он был единственной известной «полуптицей-полурептилией». Затем в отложениях мелового периода было обнаружено множество близких к археоптериксу существ, обладающих смесью признаков птиц и динозавров, которых назвали энанциорнисами (противоптицами). Оказалось, что и многие другие мелкие хищные динозавры в разное время приобретали различные птичьи черты. Найдено довольно много видов динозавров с настоящими перьями. Перья возникали либо как средство планирования при прыжках с ветки на ветку, либо для ускорения бега - у быстро бегающих наземных форм. Недавно обнаружены «четырехкрылые» динозавры, у которых перья были и на передних и на задних конечностях (рис. 13.8).
Выяснилось, что археоптерикс и родственные ему организмы предками настоящих птиц, скорее всего, не являются. Это тупиковая ветвь, вымершая в конце мелового периода и не оставившая потомков. Предками настоящих веерохвостых птиц теперь считают не пернатых динозавров, а более древних пресмыкающихся, а именно позднетриасовых текодонтов. Эта группа рептилий была общим предком и птиц, и динозавров.

Рис. 13.8. Примеры динозавров, имевших перья: а - анхиорнис; б - микрораптор
В процессе эволюции часто бывает так, что из двух параллельно развивающихся линий одна достигает успеха первой, но делает это на основе несбалансированных, несовершенных, хотя и быстро приобретенных адаптаций. Другая же, медленно развивающаяся линия, приобретает более сбалансированные адаптации и, в конце концов, побеждает. Возможно, подобная ситуация имела место и в ходе взаимоотношений неандертальцев с людьми вида Homo sapiens, которые, согласно последним данным, разошлись от общей предковой группы около 600 тыс. лет назад, а не произошли друг от друга. Очевидно, что появление человеческих черт (по аналогии с приобретением птичьих черт и т.д.) происходило параллельно в разных ветвях гоминид.
Такое параллельное приобретение признаков наблюдается не только при формировании ароморфозов, но при появлении любых других эволюционных новообразований.
Известны примеры возникновения похожих наборов жизненных форм на разобщенных континентах. Например, Южная Америка долгое время была изолирована от других материков, но еще до полной изоляции туда проникли примитивные плацентарные млекопитающие - древние копытные. Среди потомков этих животных появились организмы, поразительно схожие с настоящими лошадьми, носорогами, верблюдами - литоптерны. Причем у лошадеподобных «южных» ли-топтерн редукция пальцев ног и увеличение среднего пальца с копытом происходила практически так же, как у настоящих лошадей (рис. 13.9).

Рис. 13.9. Пример параллелизма в эволюции. Последовательные стадии филогенетических преобразований скелета стопы: I - литоптерн (а - Macrauchenia, б - Diadiaphorus, в - Thoatherium); II - лошадей (а - Eohippus; б - Miohippus; в - Meryohippus; г - Equus)
Замечательный пример параллельной эволюции - возникновение белков-антифризов, препятствующих замерзанию крови у рыб, которые живут в воде с температурой ниже точки замерзания. Такие условия существуют в Арктике и Антарктике. У антарктических и арктических рыб параллельно и независимо на разной генетической основе сформировались разные белки- антифризы.
Таким образом, накопление новых данных, применение новых методик и приемов исследования в различных областях науки позволяет расширить представление о закономерностях эволюции. Однако и сегодня эволюционистика далеко еще не разрешила всего круга стоящих перед нею проблем и продолжает свое развитие. Действие элементарных эволюционных факторов приводит к дивергенции популяций и образованию новых видов. Многообразные виды связаны друг с другом разной степенью родства. Виды, дивергировавшие от общих предков относительно недавно, сохраняют нередко значительное фено- и гено-типическое сходство. Дивергенция, произошедшая в отдаленное время, оставляет меньше общих фенотипических признаков и особенностей геномов. Виды, практически не связанные родством, характеризуются серьезными различиями.
13.1.4. Уровень организации
Говоря о прогрессивной эволюции природы в целом и животного мира в частности, следует определить понятие уровень организации. Это понятие прямо не связано со степенью сложности строения животного.
Действительно, степень сложности не только морфофизиологической организации, но и генома человека и современных человекообразных обезьян практически совпадает, но уровень организации человека благодаря его социальности, несомненно, выше.
Предложено несколько вариантов оценки уровня организации животных. Наиболее приемлемым из них является, вероятно, определение уровня организации через характер взаимоотношения организмов данной группы со средой обитания. Примитивные организмы используют во взаимоотношениях со средой так называемую стратегию r. Они ведут себя в среде пассивно, а их выживание и дальнейшая эволюция обеспечиваются в первую очередь интенсивным размножением, благодаря которому компенсируются огромные потери, возникающие в процессе естественного отбора. Для них в высшей степени характерны резкие колебания численности популяций в зависимости от сезона и изменений природноклиматических условий. В противоположность им высокоорганизованные организмы более активно проявляют себя в среде обитания и не только противостоят ей, но и способны преобразовывать ее, приспосабливая для своих нужд. Плодовитость их невелика, а выживаемость высокая, часто благодаря заботе о потомстве. Выраженных регулярных колебаний численности их популяций в естественной природе обычно не происходит. Такая стратегия взаимодействия организмов со средой носит название стратегия к. В группу организмов первого типа входят практически все беспозвоночные, в особенности, ведущие паразитический образ жизни, а из позвоночных - большинство рыб и земноводных. Стратегия К характерна для птиц и млекопитающих.
Рассмотренный критерий уровня организации можно применять только по отношению к группам организмов, связанных более или менее отдаленным родством. Так, пресмыкающиеся более высоко организованы, чем земноводные, потому, что они активнее ведут себя в среде обитания (размножение их может происходить в разнообразных условиях), при более низкой плодовитости выживаемость их потомства значительно выше. Уровень организации млекопитающих еще более высок благодаря постоянной температуре тела, сложному поведению, внутриутробному эмбриональному развитию и заботе о потомстве. Однако в одной и той же систематической группе могут находиться организмы, уровни организации которых разительно отличаются. Это касается, например, отряда Приматы, к которым относятся как низшие полуобезьяны, так и человек, уровень организации которого высок благодаря его социальному способу адаптации к среде и активному ее преобразованию (см. п. 17).
13.1.5. Направления эволюции групп
В зависимости от того, изменяется ли уровень организации в эволюционирующих группах, выделяют два основных типа эволюции: аллогенез и арогенез.
При аллогенезе у всех представителей данной группы сохраняются без изменения основные черты строения и функционирования систем органов, благодаря чему уровень организации их остается прежним. Аллогенная эволюция происходит в пределах одной адаптивной зоны - совокупности экологических ниш, различающихся в деталях, но сходных по общему направлению действия основных средовых факторов на организм данного типа. Интенсивное заселение конкретной адаптивной зоны достигается благодаря возникновению у организмов идиоадаптаций - локальных морфофизиологических приспособлений к определенным условиям существования. Пример аллогенеза с приобретением идиоадаптаций к разнообразным условиям обитания в отряде насекомоядных млекопитающих см. на рис. 13.10. Арогенез, или морфофизиологический прогресс, - такое направление эволюции, при котором у некоторых групп внутри более крупного таксона появляются новые морфофизиологические особенности, приводящие к повышению уровня их организации. Эти новые прогрессивные черты организации называют ароморфозами. Ароморфозы позволяют организмам заселять принципиально новые, более сложные адаптивные зоны. Так, арогенез ранних земноводных был обеспечен появлением у них таких основных ароморфозов, как пятипалые конечности наземного типа, легкие и два круга кровообращения с трехкамерным сердцем. Завоевание адаптивной зоны с более сложными для жизни условиями (наземной по сравнению с водной, воздушной по сравнению с наземной) сопровождается активным расселением в ней организмов с появлением у них локальных идиоадаптаций к различным экологическим нишам.
Таким образом, периоды арогенной эволюции группы могут сменяться периодами аллогенеза, когда в результате возникающих идиоадаптаций новая адаптивная зона заселяется и используется наиболее эффективно. Если в ходе филогенеза организмы осваивают более простую по сравнению с исходной адаптивную зону, например, переходя к прикрепленному неподвижному образу жизни или становясь паразитами, то они претерпевают морфофизиологический регресс, или дегенерацию, утрачивая часть прогрессивных черт, которыми обладали их предки, и оказываясь на более низком уровне организации. Нередко и новая, более простая, адаптивная зона дает возможность широких идиоадаптаций к не менее разнообразным условиям обитания в ней, т.е. обеспечивает аллогенез (рис. 13.11).

Рис. 13.10. Аллогенез в отряде насекомоядных млекопитающих. Наземные формы: а - прыгунчик; б - землеройка; в - еж; земноводные формы: г - кутора; д - выдровая землеройка; е - выхухоль; роющие формы: ж - крот; з - златокрот

Рис. 13.11. Направления эволюции групп: I - арогенез; II - аллогенез; III - дегенерация; а, б, в - адаптивные зоны
Современный органический мир характеризуется огромным многообразием форм, отличающихся как по уровню организации, так и по разнообразию локальных адаптации именно благодаря смене типов эволюции, протекающей своеобразно в каждой из эволюционирующих групп. Наряду с высокоорганизованными формами существуют примитивные, свободноживущие и паразитические организмы, возникшие эволюционно относительно недавно, и реликтовые группы, время бурного адаптационного процесса которых давно прошло.
13.1.6. Формы эволюции групп
Существуют две элементарные формы филогенеза: филетическая и дивергентная эволюция.
Филетическая эволюция - это изменения, происходящие в одном филогенетическом стволе, эволюционирующем во времени как единое целое. Реконструированы филогенетические ряды слона, лошади и других организмов, в которых можно наблюдать постепенное нарастание числа и степени выраженности признаков, характерных для современных форм (рис. 13.12).
Дивергентная эволюция заключается в образовании на основе одной предковой группы двух или нескольких производных (рис. 13.13).
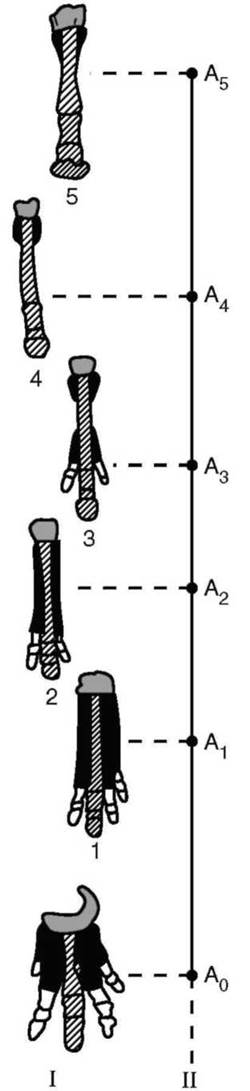
Рис. 13.12. Филетическая эволюция конечности в сем. Лошадиные: I - преобразование скелета конечности; II - схема филетической эволюции: предковая форма - фенакодус; 1 - эогиппус; 2 - миогиппус; 3 - парагиппус; 4 - плиогип-пус; 5 - лошадь; А - этапы эволюции
Она приводит к дифференциации более крупных таксонов на более мелкие, например, классов на отряды, родов на виды.
Как филетическая, так и дивергентная эволюция протекают на общей генетической базе, поэтому между организмами сохраняется более или менее выраженное генотипическое и морфофункциональное сходство.
Сопоставление филогенеза в разных группах позволяет выделить и некоторые общие закономерности соотносительной эволюции. Так, при попадании в одну и ту же среду обитания двух или более филогенетических групп неродственных организмов у них обычно проявляется конвергенция признаков. При этом сходные экологические задачи они решают сходным образом. Конвергентные адаптации возникают в этом случае на разной генетической основе, затрагивают в первую очередь поверхностные признаки, не распространяясь на общий план строения и наиболее существенные черты организации соответствующих групп. Пример конвергентной эволюции - форма тела и особенности локомоции в воде у акуловых рыб, водных пресмыкающихся - ихтиозавров, костистых рыб, пингвинов, ластоногих и китообразных млекопитающих, внутреннее строение которых полностью соответствует особенностям, характерным для классов, к которым они относятся (рис. 13.14). Конвергенция прослеживается и в эволюции глаза у позвоночных и моллюсков. Как в строении, так и в функционировании этого органа у неродственных форм обнаруживается поразительное сходство (рис. 13.15). Существенное различие глаза моллюсков и позвоночных - взаимная ориентация светочувствительных рецепторов и волокон зрительного нерва: у моллюсков рецепторы сориентированы в сторону хрусталика и зрачка, а у позвоночных они находятся на задней поверхности сетчатки. Поэтому у позвоночных место выхода зрительного нерва из глазного яблока лишено фоторецепторов и называется слепым пятном, в то время как у моллюсков его нет.

Рис. 13.13. Дивергентная эволюция клювов у вьюрковых птиц: I - возникновение разных видов в процессе изменения типов питания; II - схема дивергентной эволюции: А - D - результаты эволюции
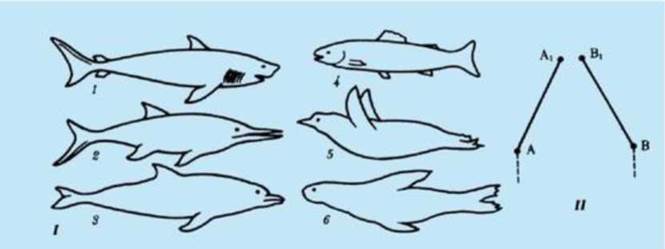
Рис. 13.14. Конвергентная эволюция формы тела позвоночных: I - водные животные разного происхождения: 1 - акула; 2 - ихтиозавр; 3 - дельфин; 4 - костистая рыба; 5 - пингвин; 6 - тюлень; II - схема конвергентной эволюции: А - Аъ В - В1 - этапы эволюции

Рис. 13.15. Конвергенция в строении и функционировании глаза у позвоночных (а) и моллюсков (б)
Другая форма соотносительной эволюции - параллелизм - реализуется в двух или нескольких группах, связанных более или менее отдаленным родством, которое основано на дивергенции от общего предка. В связи с общностью части генофондов, унаследованных от предков, у них возникают сходные адаптации в условиях действия факторов отбора в одинаковом направлении. Параллельное филетическое развитие двух родственных групп обеспечивается реализацией закона гомологических рядов (см. п. 13.3.5). Примером параллелизма является эволюция одногорбого и двугорбого верблюдов соответственно в Африке и Центральной Азии, имеющих сходные адаптации к жизни в пустынях.
Параллелизм можно наблюдать и в родственных группах организмов, разобщенных временем. Так, во второй половине кайнозойской эры в семействе кошачьих такой признак, как саблезубость (гипертрофия клыков верхней челюсти), возникал по крайней мере четырежды у представителей четырех разных родов (рис. 13.16).
13.1.7. Биологический прогресс и биологический регресс
Эволюция любого типа - арогенная, аллогенная или идущая по типу морфофизиологического регресса - в целом приводит к всесветному расселению живых организмов разного уровня организации.
Рассматривая эволюцию отдельных таксонов, можно убедиться в том, что некоторые из них находятся в состоянии расцвета, в то время как другие вымирают. Успех группы организмов в эволюционном процессе оценивают как состояние биологического прогресса, критерии которого:
• увеличение количества представителей соответствующей группы;
• расширение ареала распространения;
• активизация видоообразования в роде, увеличение количества родов в семействе, семейств в отряде и т.д.
Явление, противоположное биологическому прогрессу, - биологический регресс - свидетельствует об угасании филогенетической группы, ведущем к ее вымиранию. Так же, как и биологический прогресс, состояние регресса может длиться очень долго. Оставшиеся немногочисленные представители некогда биологически прогрессивной группы, найдя соответствующую их организации экологическую нишу, могут продолжать существовать на протяжении сотен миллионов лет. Они называются реликтами, обладают консервативностью организации и крайне медленно эволюционируют. Пример реликта - современная кистеперая рыба латимерия, ближайшие родственники которой жили в девонском периоде более 200 млн лет назад, а также дерево гинкго, непосредственные предки которого существовали на Земле уже в первой половине мезозойской эры.

Рис. 13.16. Параллелизм в эволюции саблезубости у кошачьих: I - черепа саблезубых кошек в разные периоды кайнозойской эры: а - махайрод (олигоцен - 37-25 млн лет назад); б - смилодон (миоцен - 25-9 млн лет назад); в - лжесаблезубая кошка (олигоцен - 37-25 млн лет назад); г - саблезубый тигр (плейстоцен - 1,8 млн лет назад); II - схемы параллельной эволюции: А - А2 - этапы синхронного параллелизма; В - В2 - этапы асинхронного параллелизма
На протяжении исторического развития одна и та же группа может претерпевать периоды бурного расцвета, стабильного состояния, или стагнации, и упадка. Так, в девонском периоде появились первые земноводные, вставшие на путь арогенной эволюции. Заселив наземную среду обитания, они господствовали на Земле в течение 75 млн лет. На границе палеозойской и мезозойской эр (230 млн лет назад) бурный биологический прогресс претерпевают пресмыкающиеся, быстро вытесняющие земноводных, которые с тех времен и до современного состояния переживают биологический регресс.
Расцвет пресмыкающихся длился до последней трети мезозойской эры, когда (около 65 млн лет назад) он сменился биологическим регрессом, после чего освобождающиеся вымирающими динозаврами экологические ниши стали занимать бурно эволюционирующие млекопитающие и птицы, биологический прогресс которых продолжается до настоящего времени. В то же время такой класс, как костистые рыбы, пройдя период бурного расцвета к началу мезозойской эры и прочно заняв разнообразные места обитания, до настоящего времени находится в состоянии стабильного господства среди позвоночных в водной среде обитания.
Состояние биологического прогресса может быть обусловлено как аро- и аллогенезом, так и морфофизиологическим регрессом.
Действительно, занятие новых экологических ниш как в новых адаптивных зонах, так и в уже освоенных приводит и к активизации размножения, и к расширению ареала, и к усилению видообразования.
13.1.8. Эмпирические правила эволюции групп
Из многочисленных обнаруженных эмпирическим путем правил и закономерностей эволюции групп остановимся на важнейших.
Согласно правилу необратимости эволюции, эволюция является необратимым процессом, и организмы не могут вернуться к прежнему состоянию, уже пройденному их предками ранее. Действительно, каждое эволюционно значимое изменение представляет собой комбинацию многих мутаций и появление новых регуляторных взаимодействий, подхваченных естественным отбором, но возникших случайно и независимо друг от друга. Поэтому понятно, что возвращение генофонда данной группы организмов или даже одного генотипа к состоянию, характерному для предков, статистически практически невероятно.
Так, в процессе эволюции приматов постепенно происходила утрата ими способности ощущать и дифференцировать запахи. Это было связано с тем, что по сравнению с другими млекопитающими приматы используют зрительный, слуховой и осязательный анализаторы существенно интенсивнее и чаще, чем обонятельный. В связи с этим в генах приматов, связанных с развитием и функционированием ольфакторных (обонятельных) рецепторов и обонятельного центра, произошло накопление бессмысленных мутаций, превративших эти гены в неактивные псевдогены. У человека более 70% генов этой группы представляют собой псевдогены. Естественно, что возвращение такого большого объема генетического материала к исходному состоянию невозможно.
Однако в ряде случаев за счет отдельных обратных мутаций или за счет мутаций регуляторных генов у высокоорганизованных животных возможно повторное возникновение отдельных признаков атавистической природы (см. п. 13.3.4), не меняющих в целом их тип организации.
Правило адаптивной радиации гласит, что в случае возникновения новых сред обитания, в пределах которых условия для жизни организмов оказываются достаточно благоприятными и разнообразными, эволюционный процесс ускоряется и активизируются процессы видообразования таким образом, что у организмов возникают новые идиоадаптации, обеспечивающие их эффективное выживание и дальнейший биологический прогресс. Примеры действия этого правила - возникновение многообразных видов вьюрков на Галапагосском архипелаге, адаптированных к питанию разными продуктами (см. рис. 13.13), а также занятие рыбами семейства карповых разных зон в водоеме: у дна, около поверхности, в средних слоях воды. Недостаток крупных хищных млекопитающих в начале Кайнозойской эры на территории Северной Америки и Евразии привел к тому, что их экологическую нишу заняли сухопутные крокодилы и гигантские нелетающие журавлеподобные птицы (рис. 13.17), в то время как большинство других птиц сохраняли способность к полету и адаптировались к многообразным условиям в разных природно-климатических зонах. Позже они были вытеснены более прогрессивными крупными хищными млекопитающими.

Рис. 13.17. Диатрима - нелетающая журавлеобразная хищная птица
Правило прогрессивной специализации утверждает, что филогенетическая группа, эволюционирующая по пути приспособления к данным конкретным условиям, и в дальнейшем будет продвигаться по пути углубления специализации. Генетические основы этого правила состоят в том, что в процессе направленного естественного отбора в условиях данной адаптивной зоны отсеиваются те гены генофондов популяций, которые не соответствуют ей. В результате возникает ограниченность способности генофондов к изменениям в разных направлениях.
Примером прогрессивной специализации служат морфологические преобразования конечностей в эволюционной ветви лошадей (см. рис. 13.12). При переходе к жизни на открытых пространствах с плотной почвой у предков лошади уменьшается число пальцев до одного, что не позволяет современным лошадям населять другие биотопы. Другой пример - эволюция ЮгоВосточноазиатских человекообразных обезьян гиббонов. Специализация их к древесному образу жизни при отсутствии хватательного хвоста, имеющегося, например, у всех американских древесных обезьян, привела к возникновению своеобразного типа движения - брахиации, при котором осуществляются прыжки по веткам в подвешенном состоянии на передних конечностях. Они при этом резко удлиняются, большой палец значительно редуцируется, а рука становится почти неспособной к манипуляциям мелкими предметами. При передвижении по земле руки гиббонов уже не участвуют в локомоции.
Прогрессивная специализация резко уменьшает экологическую пластичность видов и часто становится причиной их вымирания при изменившихся условиях. Уменьшение размеров и упрощение видового состава тропических лесов Индонезии - причина биологического регресса такого высокоорганизованного, но узкоспециализированного вида человекообразных обезьян, как орангутан.
Наиболее ярко выражена прогрессивная специализация в экологической группе паразитических организмов, требующих для своего существования часто абсолютно уникальных условий (см. гл. 18).
Следствие правила прогрессивной специализации - правило происхождения новых групп организмов от малоспециализированных предков. Примеров, иллюстрирующих это правило, много. Действительно, млекопитающие произошли от пресмыкающихся, сохранивших в своей организации некоторые черты земноводных. Прогрессивная линия эволюции приматов, ведущая к человеку, не связана с рамапитеком (специализированной древесной формой), как полагали раньше, а берет начало от неспециализированных африканских приматов, ведущих полуназемный образ жизни (см. п. 15.2 и 15.3). Генетическая основа этого правила - отсутствие жесткого одностороннего отбора генотипов у неспециализированных форм, сохранение в связи с этим их большего разнообразия и, следовательно, их высокой экологической пластичности.