Постоянный обмен веществ с окружающей средой - одно из основных свойств живых систем. В клетках непрерывно идут процессы биосинтеза (ассимиляция, или пластический обмен), т. е. при участии ферментов из простых органических соединений образуются сложные: из аминокислот - белки, из моносахаридов - полисахариды, из нуклеотидов - нуклеиновые кислоты и т. д. Все процессы синтеза идут с поглощением энергии. Примерно с такой же скоростью идет и расщепление сложных молекул до более простых с выделением энергии (диссимиляция, или энергетический обмен). Благодаря этим процессам сохраняется относительное постоянство состава клеток. Синтезированные вещества используются для построения клеток и их органоидов и замены израсходованных или разрушенных молекул. При расщеплении высокомолекулярных соединений до более простых выделяется энергия, необходимая для реакций биосинтеза.
Совокупность реакций ассимиляции и диссимиляции, лежащих в основе жизнедеятельности организма и обусловливающих его связь с окружающей средой, называется обменом веществ.
Для реакций обмена характерна высокая организованность и упорядоченность. Каждая реакция протекает с участием специфических белков-ферментов. Они располагаются в основном на мембранах органоидов и в гиалоплазме клеток в строго определенном порядке, что обеспечивает необходимую последовательность реакций. Благодаря ферментным системам реакции обмена идут быстро и эффективно в обычных условиях - при температуре тела и нормальном давлении.
Пластический и энергетический обмены неразрывно связаны. Они являются противоположными сторонами единого процесса обмена веществ. Реакции биосинтеза нуждаются в энергии, которая поставляется реакциями энергетического обмена. Для осуществления реакций энергетического обмена необходим постоянный биосинтез ферментов и структур органоидов, которые в процессе жизнедеятельности постепенно разрушаются.
Процессы ассимиляции не всегда находятся в равновесии с процессами диссимиляции. Так, в растущем организме процессы ассимиляции преобладают над процессами диссимиляции, благодаря чему обеспечивается накопление веществ и рост организма. При интенсивной физической работе и в старости преобладают процессы диссимиляции. В первом случае это компенсируется усиленным питанием, а во втором происходит постепенное истощение и в конечном итоге гибель организма.
Пластический обмен (анаболизм) - совокупность реакций биологического синтеза, при котором из поступивших в клетку веществ образуются вещества, специфические для данной клетки.
К пластическому обмену относятся фотосинтез, биосинтез белков, синтез нуклеиновых кислот, жиров и углеводов.
По типу питания живые организмы делятся на две группы - автотрофные и гетеротрофные.
Гетеротрофными называются организмы, не способные синтезировать органические вещества из неорганических и использующие в качестве пищи (источника энергии) готовые органические соединения. К гетеротрофам относятся большинство бактерий и протистов, грибы и животные.
Автотрофными называются организмы, способные синтезировать из неорганических веществ органические, служащие строительным материалом и источником энергии. К ним относятся пигментированные бактерии, некоторые протисты и все зеленые растения. Автотрофные организмы подразделяются на хемосинтезирующие и фотосинтезирующие. Хемосинтезирующие организмы (бактерии) потребляют энергию, выделяющуюся при окислении некоторых неорганических веществ (например, нитрифицирующие бактерии последовательно окисляют аммиак до нитритов, а затем нитриты до нитратов).
Фотосинтезирующие организмы (зеленые растения) используют энергию света.
Зеленые растения способны с помощью пигментов хлорофиллов и каротиноидов, содержащихся в хлоропластах, преобразовывать световую энергию Солнца в энергию химических связей органических веществ. В частности, из энергетически бедных веществ СО2 и Н2О они синтезируют богатые энергией углеводы и выделяют кислород. Этот процесс синтеза первичного органического вещества называется фотосинтезом, который представляет собой цепь окислительно-восстановительных реакций. Он протекает в две фазы: световую и темновую (рис. 114).
Фотосинтез начинается с момента освещения хлоропласта видимым светом. Улавливание солнечной энергии происходит светочувствительными пигментами, которые образуют светособирающие комплексы, работающие наподобие воронки. Далее световая энергия передается от молекулы к молекуле и в конечном итоге доставляется на специализированную форму хлорофилла. Совокупность молекул хлорофилла и ферментов, обеспечивающих транспорт электронов, называется фотосистемой. Различают два типа фотосистем. Фотосистема I, содержащая хлорофилл с максимальным поглощением света (длина волны 700 нм) - Хл700, и вторая - с максимальным поглощением света (длина волны 680 нм) - Хл680.
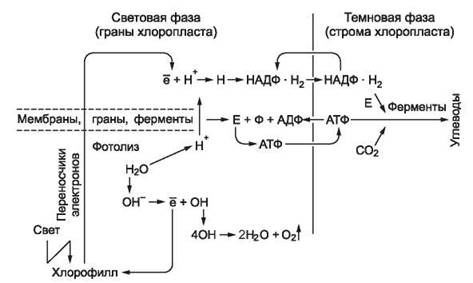
Рис. 114. Схема фотосинтеза
При поглощении молекулами хлорофилла фотосистемы I квантов света их электроны переходят в «возбужденное» состояние, поднимаются на более высокий энергетический уровень, поступают к акцептору и далее в цепь переноса электронов, а затем переходят на наружную поверхность мембран гран. При этом Хл700 окисляется и превращается в Хл+700. Возбужденные молекулы хлорофилла фотосистемы II (Хл680) отдают свои электроны акцептору. По цепи переноса электронов они поступают в фотосистему I и восстанавливают молекулы Хл+700 до Хл700. Молекула восстановленного хлорофилла (Хл700) вновь способна переходить в возбужденное состояние. Молекулы Хл680, отдав свои электроны, окисляются до Хл+680, восстановление которых происходит за счет электронов, образующихся при фотолизе воды. Под действием света вода распадается на ионы Н+ и ОН-. Протоны накапливаются на внутренней поверхности мембран гран хлоропласта (электроны - на наружной) и создают электрохимический мембранный потенциал. Когда разность потенциалов достигает критического уровня (около 200 мВ), протоны проходят по специальным каналам мембран, в которых находятся ферменты АТФ-синтетазы. Энергия протонов и электронов используется ферментами для присоединения остатков фосфорной кислоты (фосфорилирование) к АМФ или АДФ. После прохождения по каналам протоны взаимодействуют с электронами и никотинамидаденинди- нуклеотидфосфатом (НАДФ) и восстанавливают его до НАДФН+Н+. Ионы гидроксила, оставшись без противоионов Н+, отдают свои электроны и превращаются в свободные радикалы ОН, которые, взаимодействуя друг с другом, образуют воду и свободный кислород:
4ОН → 2Н2О + О2.
Электроны гидроксильных групп идут на восстановление Хл+680. Таким образом, в световую фазу фотосинтеза, которая протекает в гранах хлоропластов только на свету, происходят следующие процессы: фотолиз воды с выделением кислорода, восстановление НАДФ до НАДФН+Н+ и синтез АТФ.
В темновую фазу фотосинтеза накопленная в световую фазу энергия используется для синтеза моносахаридов из диоксида углерода (поступает из воздуха через устьица) и водорода (отсоединяется от НАДФ Н + Н+) путем сложных многоступенчатых ферментативных реакций (цикл Кальвина). В итоге получается:
6СО2 + 24Н → С6Н12О6 + 6Н2О.
В дальнейшем могут образовываться ди-, полисахариды и другие органические соединения (аминокислоты, жирные кислоты и др.). Этот процесс не требует прямого участия света. Он протекает в строме хлоропластов как на свету, так и в темноте. Коэффициент полезного действия фотосинтеза достигает 60%. Фотосинтез и дыхание являются противоположными процессами, их сравнительная характеристика дана в табл. 10.
Таблица 10
Сравнительная характеристика процессов фотосинтеза и дыхания
|
Фотосинтез |
Дыхание |
|
Образование органических веществ Поглощение СО2 Выделение О2 Накопление энергии Происходит только на свету Происходит в клетках, содержащих хлорофилл |
Распад органических веществ Поглощение О2 Выделение СО2 Освобождение энергии Происходит на свету и в темноте Происходит во всех живых клетках |
Значение фотосинтеза огромно. Это главный процесс синтеза первичного органического вещества, протекающий в биосфере. Энергия Солнца аккумулируется в химических связях органических соединений, которые идут на питание всех гетеротрофов. При этом атмосфера обогащается кислородом и очищается от избытка диоксида углерода.
Биосинтез белков осуществляется во всех клетках про- и эукариотических организмов, это неотъемлемое свойство живого. Информация о первичной структуре белковой молекулы, от которой зависят все остальные структуры и свойства, закодирована последовательностью нуклеотидов в соответствующем участке молекулы ДНК - гене. Так как информация о структуре молекулы белка находится в ядре, а его сборка идет в цитоплазме (в рибосомах), в клетке имеется посредник, копирующий и передающий эту информацию. Таким посредником является информационная РНК (и-РНК). Специальный фермент (РНК-полимераза) расщепляет двойную цепочку ДНК, и на одной из ее цепей (кодирующей) по принципу комплементарности выстраиваются нуклеотиды РНК. Считывание информации с кодирующей цепочки ДНК идет в направлении 3' →5'. Таким образом, синтезированная молекула и-РНК повторяет порядок нуклеотидов в ДНК. Этот процесс называется транскрипцией (переписыванием). Синтезированная таким образом (матричный синтез) молекула и-РНК выходит в цитоплазму и на один ее конец (5 ) сначала нанизывается малая субъединица рибосомы, а затем присоединяется большая субъединица (происходит сборка рибосомы), что обеспечивается инициирующим кодоном (АУГ).
Система записи генетической информации в ДНК (и-РНК) в виде определенной последовательности нуклеотидов называется генетическим кодом.
Свойства генетического кода:
+ триплетность - одной аминокислоте в полипептидной цепочке соответствуют три расположенных рядом нуклеотида молекулы ДНК (и-РНК), называемые триплетом, или кодоном;
+ универсальность - одинаковые кодоны кодируют одну и ту же аминокислоту у всех живых организмов;
+ неперекрываемость - один нуклеотид не может входить одновременно в состав нескольких кодонов;
+ избыточность (вырожденность ) - одну аминокислоту могут кодировать несколько различных триплетов;
+ отсутствие разделительных знаков внутри гена при их наличии между генами (начало считывания информации с и-РНК определяется инициирующим кодоном - АУГ, а конец - терминирующими кодонами - УАА, УАГ, УГА).
Следующий этап в биосинтезе белка - перевод последовательности нуклеотидов в молекуле и-РНК в последовательность аминокислот в полипептидной цепочке - трансляция. Транспортные РНК (т-РНК) «приносят» аминокислоты в рибосому. Молекула т-РНК имеет сложную конфигурацию (рис. 115). На некоторых участках ее между комплементарными нуклеотидами образуются водородные связи, и молекула по форме становится похожей на лист клевера. На его «верхушке» расположен триплет свободных нуклеотидов, которые по своему генетическому коду соответствуют данной аминокислоте (он называется антикодоном), а «черешок» (основание) служит местом прикрепления этой аминокислоты. Каждая т-РНК может переносить только свою аминокислоту.
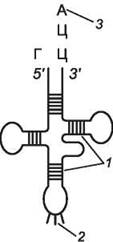
Рис. 115. Схема структуры молекулы т-РНК: 1 - водородные связи; 2 - антикодон; 3 - место прикрепления аминокислоты
На одном конце (5') молекулы т-РНК («черешка») всегда находится гуанин, а на другом (3') - триплет ЦЦА. Каждая аминокислота присоединяется к одной из своих т-РНК при участии специфического фермента (аминоацил-m-РНК-синтетазы) и АТФ (активация аминокислот). В результате образуется комплекс аминокислоты с т-РНК (аминоацил-m-РНК), в котором энергии связи между концевым нуклеотидом А (в триплете ЦЦА) и аминокислотой достаточно для образования в дальнейшем пептидной связи. Т-РНК с аминокислотой поступает в аминоацильный (аминокислотный) центр рибосомы (рис. 116). Если антикодон т-РНК является комплементарным кодону и-РНК, то происходит временное присоединение т-РНК с аминокислотой к и-РНК. Рибосома передвигается по и-РНК на один кодон влево. Т-РНК с аминокислотой, находившаяся в аминоацильном центре, оказывается в пептидильном центре. В свободный аминоацильный центр поступает следующая т-РНК со своей аминокислотой и, если антикодон и кодон комплементарны, присоединяется к и-РНК. В рибосоме в каждый данный момент находятся два кодона и-РНК. Аминокислоты располагаются у активного центра большой субъединицы рибосомы, и с помощью ферментов между ними устанавливается пептидная связь. Одновременно разрушается связь между первой аминокислотой и ее т-РНК; т-РНК отсоединяется от и-РНК и уходит из рибосомы за следующей аминокислотой. Рибосома перемещается на один триплет влево, и процесс повторяется. Так постепенно наращивается молекула полипептида, в которой аминокислоты располагаются в строгом соответствии с порядком кодирующих их триплетов (матричный синтез). Конец считывания информации определяется кодонами-терминаторами. Часто на одну и-РНК нанизывается не одна рибосома, а несколько (такие структуры называются полисомами); при этом синтезируется несколько одинаковых белковых молекул.
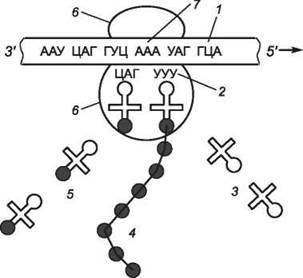
Рис. 116. Схема биосинтеза белка: 1 - и-РНК; 2 - антикодон; 3 - т-РНК; 4 - полипептидная цепочка; 5 - т-РНК с аминокислотами; 6 - субъединицы рибосом; 7 - триплет
После завершения синтеза белковая молекула отделяется от рибосомы и приобретает свойственную ей (вторичную, третичную или четвертичную) структуру. Биосинтез белка идет довольно быстро. За 1 с бактериальная рибосома образует полипептид из 20 аминокислот. Скорость биосинтеза зависит от активности ферментов, катализирующих процессы транскрипции и трансляции, от температуры, концентрации водородных ионов, наличия АТФ и свободных аминокислот и др.